√лава 4. ƒонные, придонные и придонно-пелагические рыбы
»зучение рыб только по коллекционным образцам и так называемым Ђполевым биологическим анализамї (длина и масса тела, врелость гонад, наполнение желудков и т. п.) пикак по может считатьс€ полноценным - оно почти не дает представлени€ ни об их образе жизни, ни, тем более, поведении. Ёти аспекты биологии особенно важно знать, когда имеешь дело с донно-придонными рыбами,- ведь они отличаютс€ исключительным разнообразием взаимоотношений и поведенческих реакций. ≈сли еще учесть, что рыб, подн€тых с глубины, трудно содержать в аквариумах, становитс€ пон€тным, почему об их поведении до сих пор известно так мало. ѕочитайте, к примеру, Ђ∆изнь животныхї Ч соответствующие разделы часто содержат лишь описани€ внешнего вида и сведени€ о глубине и месте поимки... ћежду тем в последние годы по€вилась прекрасна€ возможность преодолеть эти затруднени€ Ч она св€зана с использованием автономных обитаемых подводных аппаратов.
“акие аппараты есть и в »нституте океанологии, и два из них (они рассчитаны на глубину до 2000 м) даже нос€т вполне ихтиологическое название ЂPiscesї (по-латински Ђрыбыї). превеликому сожалению, ни €, ни кто-либо из коллег-ихтиологов на этих аппаратах-рыбах не погружалс€ (в основном они обслуживают геологов), а наблюдени€, сделанные нашими Ђштатными подводникамиї, надо пр€мо сказать, большой научной ценности не представл€ют (трудно поверить на слово, например, рассказу о том, что в расном море не то винцигуерии, не то миктофиды зарываютс€ в грунт на большой глубине). онечно, фотографии и видеозаписи более информативны, но без осмысленного выбора нужного ракурса они далеко не всегда позвол€ют точно определить запечатленный объект.
¬ то же врем€ другими исследовател€ми уже выполнено немало ценных наблюдений над поведением глубоководных рыб. ѕриведу несколько примеров. ѕогружа€сь в 1975 г. на аппарате ЂЁдвинї на материковом склоне северо-восточного побережь€ —Ўј, ƒэн оэн определил скорость антиморы, уходившей от преследовани€ аппарата (39,2 см/с), и рассчитал, что расход энергии на плавание (3,6.105 эрг/с) у нее примерно одинаков с форелью того же размера - рыбой, известной своими скоростными качествами, ¬ 1977 г. он же впервые сфотографировал па глубине около 2000 м самку речного угр€, пришедшую на нерест к Ѕагамским островам.—евастопольские ихтиологи √еоргий √оловапь и Ќиколай ѕахоруков в 1979 г. проследили с аппарата Ђ“»Ќ–ќї, как на одной из гор подводного хребта Ќаска в ёго-¬осточной ѕацифике беспузырные окуни вместе с лангустами уходили с плоской вершины гайота на его склоны при подтоке с севера обедненной кислородом воды и снова по€вл€лись па ней, когда ситуаци€ мен€лась и на вершину вновь поступала вода, богата€ кислородом.
...я продолжаю наде€тьс€, что и мне доведетс€ когда-нибудь увидеть глубоководных рыб в естественной дл€ них обстановке, ј пока € наблюдал их в лучшем случае полуживыми - умирающими после изъ€ти€ из родной стихии.
—огласно прин€тому в этой книге определению Ђоткрытого океанаї счет его донно-придонным биотопам начинаетс€ с материкового склона. –ассматрива€ субконтинентальный бентопелагический комплекс ихтиоценов, нужно иметь в виду, что в св€зи с сильной расчлененностью склона, преп€тствующей успешным тралени€м, население мезо- и батибентали изучено значительно хуже, чем сообщества всех прочих крупномасштабных биотопов. “ем не менее изучение батиальных рыб в последние годы значительно продвинулось.
ѕрежде всего следует отметить, что в самых верхних горизонтах мезобентали насто€щие среднеглубинные рыбы сосуществуют с сублиторальными, примесь которых в ихтиоцене может быть очень значительной. Ёта переходна€ зона должна рассматриватьс€ поэтому как экотон. ¬близи 200-метровой изобаты виды, общие дл€ эпи- и мезобентали, настолько многочисленны, что нет смысла перечисл€ть их здесь. Ќекоторые эврибатные формы встречаютс€ в очень широком диапазоне глубин. ћногие из них представлены у нижних пределов своего вертикального распространени€ более крупными особ€ми, чем на мелководь€х, дл€ других, имеющих широкие ареалы, характерно увеличение глубины обитани€ в тропической зоне, но в целом мелководные и среднеглубинные виды обычно неплохо различаютс€ по предпочитаемой глубине. ѕоэтому обыкновенного катрана (Squalus acanthias), например, наиболее обычного на глубине 20Ч200 м, относ€т к сублиторальным видам, а черного палтуса (Reinhardtius hippoglossoides), во взрослом состо€нии концентрирующегос€ на 150Ч800 м, Ч к мезобентальным.
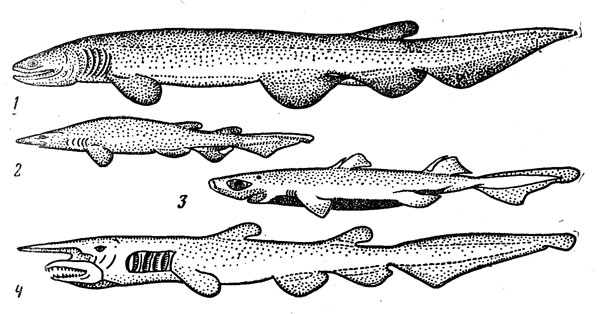
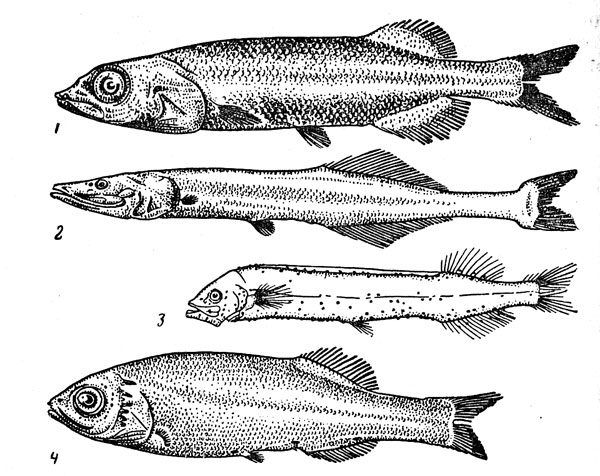
–ис. 31. ѕридонные акулы: плащеносна€ (1), глубоководна€ кошачь€ (2), этмоптерус (3), акула-домовой (4).
ћезобентопелагический комплекс ихтиоценов исключительно разнообразен по составу. —реди характерных дл€ него щележаберных рыб нужно отметить плащеносных (сем. Chlamydoselachidae), колючих (Squalidae) и кошачьих (Scyliorhinidae) акул, а также ромбовых скатов (Rajidae).
ќчень своеобразна€ плащеносна€ акула (Chlamydoselachus anguineus) (рис. 31) Ч представитель одной из наиболее примитивных групп щележаберных Ч имеет угревидно удлиненное тело и жаберные щели, прикрытые кожистыми складками (их по шесть с каждой стороны вместо п€ти у большинства акул). ќна может достигать 2 м в длину, но половозрелость наступает при значительно меньших размерах Ч у самцов около 1 м, самок Ч 1,3 м, –азмножаетс€ путем €йцеживорождени€, принос€ от 8 до 12 детенышей длиной 40 см, причем считаетс€, что вынашивание эмбрионов продолжаетс€ очень долго (год или даже два). Ёта редка€ акула обитает на материковом склоне и подводных подн€ти€х (глубина ловов варьирует от 120 до 1300 м) в умеренно теплых и субтропических –айонах всех океанов. Ѕлизкие к ней и тоже не очень часто встречающиес€ семи- и шестижаберные акулы (рода Heptranchias и Hexanchus) также достигают довольно крупных размеров (1,5Ч4,8 м) и ведут придонный образ жизни в широком диапазоне глубин шельфа и склона.
олючие акулы, напротив, принадлежат к числу самых обычных обитателей мезобентопелагиали Ч в ее пределах встречаютс€ 58 из 68 видов семейства (еще 10, как уже говорилось, живут в пелагиали без св€зи с дном), причем 46 видов обитают только в этом биотопе, а три €вл€ютс€ общими дл€ мезо- и батибентопелагического комплекса ќдин из них встречаетс€ и над ложем океана Ч португальской акуле Centroscymnus coelolepis, поимка которой зарегистрирована в 3675 м от поверхности, принадлежит рекордное по глубине нахождение среди всех хр€щевых рыб (правда, еще глубже Ч па глубине более 6000 м наблюдали из батискафа не определенных до вида колючих акул, отнесенных к роду Centrophorus). —овокупный ареал семейства простираетс€ от јрктики до —убантарктики. ќчень широко распространены и многие виды, обнаруженные во всех трех океанах: эврибатный обыкновенвый катран, например, обитает в умеренных водах обоих полушарий, а в тропиках его почти повсюду замещает малошипый катран (Squalus mitsukurii).
олючие акулы сильно различаютс€ по размерам. ƒлина тела у мелких видов рода Etmopterus не превышает 25Ч30 см (к этому роду относ€тс€ самые мелкие из ныне живущих акул: ≈. carteri и ≈. perryi станов€тс€ половозрелыми при длине 16Ч18 см и вырастают всего лишь да 21 см), а у гигантских представителей рода Somniosia достигает 6 м и более. Ќаибольша€ из достоверно измерена ных пол€рных акул (S. microcephalus) имела 6,4 м в длину при массе около 1 т, но есть сообщени€ и о еще более крупных экземпл€рах ( . √. онстантинов отмечал поимку в 1956 г. двух 8-метровых особей у берегов ћурмана) Ётот вид распространен в северной јтлантике (на юг до залива ћэн и ѕиренейского полуострова), а также у ёна ной јфрики и субантарктических островов. ¬ высоких широтах пол€рна€ акула нередко (особенно зимой) встречаетс€ на мелководь€х Ч у √ренландии эскимосы промышл€ют ее через проруби в припае, а в умеренно теплых водах предпочитает глубины склона (до 1200м). ќна питаетс€ разнообразной рыбой, часто нападает на тюленей может пожирать любые отбросы и падаль. ƒолгое врем€ считалось доказанным, что эта акула в отличие от вс€ других представителей семейства размножаетс€, откладыва€ в воду м€гкие €йца (это представление зафиксирован во всех учебниках ихтиологии), но в действительности oна €йцеживород€ща, как и все ее родичи: из самки длиной около 5 м было извлечено 10 готовых к рождению эмбрионов длиной 37 см.
Ќекоторые колючие акулы (в том числе обыкновенные катран) ведут стайный образ жизни и совершают значительные по прот€женности сезонные миграции. »нтересно, что собравшиес€ в стаю особи одного из самых мелких видов Ч карибско-мексиканского Etmopterus virens, живущего на глубине 350Ч465 м, могут совместно атаковать крупную добычу, в частности больших кальмаров, и разрывать ее на куски. Ёти акулы, подобно многим другим сквалидам, имеют фотофоры и обладают способностью к свечению. –ост колючих акул практически не изучен. ƒанные по возрасту имеютс€ только дл€ обыкновенного катрана, который достигает зрелости на 10Ч20-м году и живет по меньшей мере до 25Ч30 лет. ѕлодовитость у разных видов варьирует от 2 до 20 эмбрионов.
Ѕольшое семейство кошачьих акул, включающее около 90 видов, также очень характерно дл€ субконтипентальной мезобентали, в которой они представлены 62 видами (остальные обитают в прибрежной зоне). Ќа значительных глубинах особенно обычны представители рода Apristurus, из которых четыре даже встречены глубже 1500 м (A. microps добывалс€ только в диапазоне глубин от 1000 до 2000 м). ¬се кошачьи акулы ведут оседлый образ жизни и не совершают больших перемещений, а их apeaлы, как правило, невелики по площади. ќни имеют небольшие размеры: многие виды достигают половой зрелости уже при длине 20 см, а самые крупные из среднеглубинных форм Ч проникающие с шельфа на склон акулы-кошки рода Scyliorhinus Ч не превышают 1,2 м в длину.
¬ этом семействе наблюдаетс€ значительное разнообразие типов размножени€. Ќаиболее примитивный способ состоит в последовательной откладке на субстрат каждого оплодотворенного €йца, заключенного в прочную капсулу, котора€ прикрепл€етс€ ко дну роговыми нит€ми; развитие зародыша продолжаетс€ без вс€кой охраны в течение продолжительного времени Ч около года. ” других видов €йца упаковываютс€ в капсулы по нескольку штук и про ход€т начальные этапы развити€ в €йцеводах матери, так что врем€ между откладкой капсулы на дно и вылуплением детенышей сокращаетс€ до нескольких недель. ≈сть реди кошачьих акул и €йцеживород€щие виды, у которых эмбрионы остаютс€ в €йцеводах вплоть до полного рассеивани€ желточного мешка и рождаютс€ на свет вполне товыми к самосто€тельному существованию. ѕримеры всех трех способов размножени€ можно найти, в частности ” разных видов из родов Halaelurus или Galeus. “ак, энский галеус (G. nipponensis) откладывает одиночные €йца, чернорота€ акула (G, melastomus), обычна€ на материковом склоне ≈вропы, вынашивает в €йцеводах до 13 €иц, упакованных в капсулы дл€ последующей откладки на дно, а западноафриканский вид G. polli способен в €йцеживорождепию. ћало того, ‘. ‘. Ћитвинов сообщил мне, что он наблюдал у последнего вида даже внутриутробное поедание неоплодотворенных €иц, которое считалось до сих пор свойственным только ламноидным акулам (об этом уже говорилось в гл. II). ¬ эмбриональной оофагии и каннибализме подозревалась, правда, и ложнокунь€ акуле (Pseudotriakis microdon) Ч довольно крупный (длина до 3 м) мезобентальный вид, известный с глубины 200-1500 м во всех океанах. Ёта акула приносит всего 2Ч4, но весьма солидных по размерам (70Ч85 см) детенышей, нар€ду с которыми в €йцеводах находили большие запасы желтковых €иц (до 20 тыс. в каждом €йцеводе у самки длиной 2,8 м); это подозрение остаетс€, однако, недоказанным, так как желтка в желудках эмбрионов пока не обнаружено.
»з других глубоководных акул нужно прежде всего упом€нуть митсукурину, или акулу-домового (Mitsukurinai owstoni), выдел€ющуюс€ своей необычной, пр€мо-таки неправдоподобной внешностью: ее сильно удлиненное рыло имеет форму заостренного сошника, а челюсти обладают способностью выдвигатьс€ далеко вперед, что св€зано по-видимому, с питанием донными животными. Ёта акула известна€ по считанным поимкам, достигает 4 м в длину. ќна ловилась на глубине до 550 м, но обломанный зуб, несомненно, принадлежащий этому виду, был извлечен однажды из пробитого телеграфного кабел€, проложенного по дну в 1350 м от поверхности. ѕара митсукурин была доставлена в јтлантический институт рыбного хоз€йства и океанографии в алининграде, и чучело одной из них украшает сейчас музей этого учреждени€.
ќтдельные виды, вход€щие в глубоководные бентопелагические комплексы, есть и в некоторых преимущественно шельфовых семействах акул. —реди куньих акул (сем. Triakidae), например, один вид (Jago omanensis) известен по левам на глубине 110Ч2195 м. “олько в водам свала глубин (600Ч1000 м) обитает большеглаза€ песчана€ акула Odontaspis noronhai. ѕридонный вид, т€готеющий к глубинам, есть даже среди серых акул: большеноса€ акула (Carcharhinus altimus), встреченна€ во всех океанах придерживаетс€ главным образом переходной зоны между эпи- и мезобенталью (обычна на глубине 90Ч430 м). ѕо внешним признакам она очень близка к одному из прибрежных видов (—. plumbeus), от которого отличаетс€ лишь мелкими детал€ми строени€.
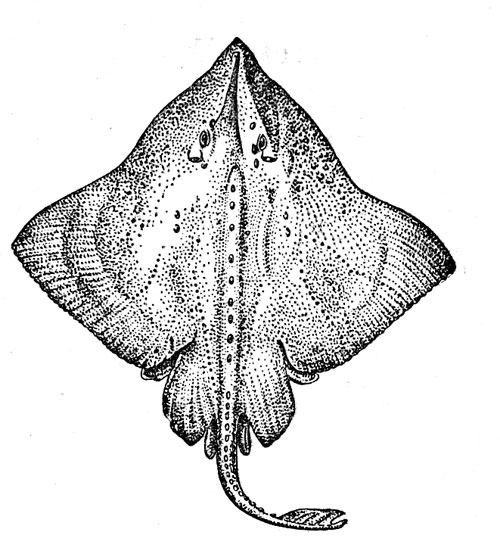
–ис. 32. —еверный ромбовый скат.
–омбовые скаты (рисунок 32) Ч типично донные рыбы, ќни провод€т свою жизнь, лежа на грунте, в который могут закапыватьс€ так, что на поверхности остаютс€ только глаза, брызгальца и часть спины, или плава€ в придонном слое. »х пищу составл€ют различные рыбы и беспозвоночные (преимущественно ракообразные), причем при охоте на плавающую добычу скаты наплывают на жертву сверху и прижимают ее ко дну своим диском, а потом уже заглатывают. ¬ этом семействе, систематику которого интенсивно разрабатывают сейчас специалисты разных стран Ч ћ. Ўтеманн из √амбурга, владивостокский ихтиолог ¬. Ќ. ƒолганов, €понец X. »сихара и др., еще недавно числили 170Ч180 видов, а теперь их насчитывают уже более 200 (почти половина всего подотр€да скатов), однако и эта цифра будет, без сомнени€, увеличена. –омбовые скаты живут во всех океанах и мор€х Ч их ареал простираетс€ от јрктики до јнтарктики. ќни особенно характерны дл€ холодных и умеренно теплых районов, но встречаютс€ и в тропической зоне, где обитают обычно на значительной глубине. Ѕольша€ часть видов (в том числе почти все представители самого обширного рода Raja, включающего около 140 видов) имеет центром вертикального диапазона распределени€ зону, расположенную на границе шельфа и склона, и относитс€, таким образом, к эпимезобентальной группировке. Ќаиболее глубоководные формы, которые принадлежат большей частью к родам Bathyraja (это крупные скаты длиной около 2 м) и Breviraja (их длина 30Ч40 см), вполне обычны и в батибентали: на рекордной глубине 2400Ч2950 м выловлены в Ѕискайском заливе типовые экземпл€ры Bathyraja pallida. –омбовые скаты размножаютс€, откладыва€ на дно €йца, заключенные по одному в роговые капсулы с характерными трубчатыми отростками по углам. ак показали наблюдени€ в аквариуме, самка обычно выметывает €йца парами с перерывом в 1Ч5 дней, и общий период размножени€ может быть очень длительным. „исло отложенных капсул, по-видимому, сильно варьирует у разных видов: от 4Ч5 до 50 и более. »нкубаци€ €иц длитс€ от 4 до 14 мес€цев. —каты принадлежат к долгоживущим видам. ѕо данным ¬. Ќ. ƒолганова, батиальные виды, обитающие в дальневосточных мор€х, достигают половой зрелости в возрасте 3Ч10 лет, а живут от 7 до 18 лет (в зависимости от размера вида).
¬не шельфов представлены и некоторые другие группы скатов, в том числе три тропических семейства, близкородственные ромбовым скатам Ч Anacanthobatidae (нитерылые скаты), Arhynchobatidae (безрылые) и Pseudorajidae (ложноромбовые), которые в полном составе принадлежат к мезобентальиому ихтиоцену. нему же относ€тс€ очень своеобразные шестижаберные скаты (сем. Ќехаtrygonidae) Ч хвостоколы с шестью жаберными щел€ми. ѕервый их представитель Ч Hexatrygon bickelli Ч был найден в 1981 г. выброшенным на берег у ѕорт-Ёлизабет (ёжна€ јфрика); в том же году другой вид этого рода был описан по 2 экз. из ёжно- итайского мор€, а затем к ним добавилась пара рыб, пойманных у островов –юкю и √авайских в диапазоне глубин от 350 до 1000 м. ≈ще один молодой самец, выловленный в »ндийском океане вблизи острова ява, был обнаружен недавно ё. Ќ. ўербачевым среди не определенных рыб в коллекции «оологического музе€ ћ√”.
Ќа довольно больших глубинах обитают и некоторые электрические скаты из семейства гнюсовых (Torpedinidae), обладающие, как хорошо известно, своеобразными электрическими органами, расположенными по бокам тела между головой и грудными плавниками. “ак, многие виды рода Torpedo, обычные на шельфе, могут встречатьс€ вплоть до глубины 400Ч500 м, а “. tokionis из вод японии и индийский Benthobatis moresbyi (сем. Narkidae) живут только на склоне (220Ч1100 м). ак это ни странно, некоторые гнюсы (например, “. tremens из юго-восточной ѕацифики) способны и к пелагическому образу жизни; этот факт вызывает удивление, поскольку все электрические скаты плохие пловцы: их единственным движителем служит короткий хвостовой стебель с небольшой лопастью хвостового плавника.
огда мне впервые довелось увидеть живого гпюса в траловом улове (это было на горе Ѕольша€ - одном из гайотов подводного хребта Ќаска), € недолго боролс€ с искушением испытать на себе действие его электрического разр€да. ќсобого геройства в этом, конечно, не было - ведь во всех сводках и руководствах вполне четко сказано, что максимум напр€жени€ Ђбатарейї этих рыб не превышает бытовых 220 вольт (€ и сам писал об этом в Ђ∆изни животныхї). “ем не менее € вз€л ската в руки внутренне готовый к тому, что сейчас ощутимо Ђдернетї, но... √нюс лишь в€ло шевельнул хвостом, €вно не жела€ показывать свое умение. я пом€л ему брюхо и спину, пощекотал пальцем голову. Ќикакого эффекта. ќтложив ската в сторону, € стал выбирать из улова, надо сказать довольно скудного, других рыб, заслуживавших внимани€, и этот отбор зан€л минут дес€ть. —обравшись в лабораторию, € вз€л ведро с рыбой и небрежно ухватил гнюса за хвост, на что он тут же ответил довольно чувствительным (жаль, не было вольтметра) и совершенно неожиданным разр€дом, на который € отреагировал не менее быстро, но, к сожалению, отнюдь не лучшим и, главное, столь же неожиданным дл€ самого себ€ образом Ч одним движением выбросил обидчика за борт.
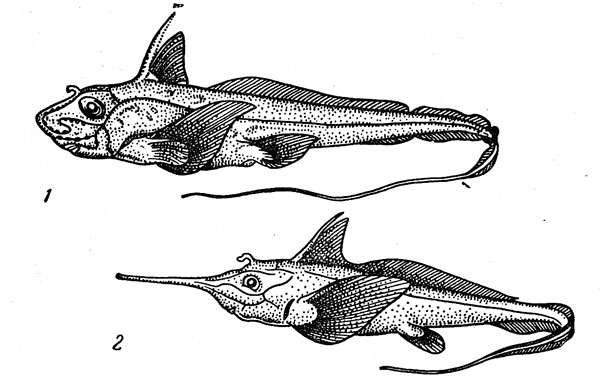
–ис. 33. ≈вропейска€ химера (1) и ринохимера (2).
ƒать правдоподобное объ€снение происшедшему не так уж трудно: по-видимому, электрический скат еще в трале израсходовал всю емкость своих батарей и на подзар€дку севших элементов потребовалось врем€, которого как раз хватило до моего вторичного прикосновени€.
ак следует из сказанного выше, доминирующие в мезобентопелагических комплексах группы щележаберных рыб существенно различаютс€ по тесноте св€зи с твердым субстратом Ч ромбовые скаты, провод€щие большую часть времени непосредственно на дне, могут быть отнесены (с очень небольшой нат€жкой) к бентосу, кошачьи акулы, также нередко поко€щиес€ на грунте, Ч к нектобентосу, а колючие акулы Ч к бентонектону. —оответственно этому колючие акулы значительно чаще встречаютс€ в пелагиали (некоторые из них даже над большими глубинами), чем представители двух других семейств. ѕространственное распределение этих рыб отражает различи€ их экологии: переход от донного образа жизни к придонно-пелагическому сопровождаетс€ изменением типа размножени€ (от откладки капсулированных €иц к €йцеживорождению) и состава пищи (от потреблени€ бентических беспозвоночных к охоте на рыб и кальмаров в водной толще).
≈ще одна очень характерна€ группа глубоководных донных рыб Ч цельноголовые, которые представлены в мезо- и батибентали двум€ семействами (рис. 33). ’имеровые (Ghimaeridae) обитают преимущественно в верхней части материкового склона на глубине от 200 до 700Ч 1000 м. »зученна€ лучше других европейска€ химера (Chimaera monstrosa), обитающа€ в восточной јтлантике (один экземпл€р был пойман в 1957 г. в советских водах Ч у берегов восточного ћурмана), достигает 1,5 м в длину. ƒругие представители семейства (в нем числ€тс€ два рода и более 20 видов), как правило, также имеют довольно ограниченное распространение, хот€ общий его ареал охватывает батиаль всех тропических и умеренно теплых районов ћирового океана, причем у краев этой широтной зоны химеры встречаютс€ и на относительно небольших глубинах. Ѕудучи медлительными и неповоротливыми, эти рыбы посто€нно держатс€ у дпа. ќни плохие пловцы, передвигающиес€ только использу€ в качестве весел свои вееровидные грудные плавники. ѕищу химер составл€ют донные беспозвоночные и рыбы, а размножаютс€ они, откладыва€ на дно веретеновидные €йцевые капсулы, которые прикрепл€ютс€ к субстрату специальными роговыми нит€ми.
Ѕлизкое семейство носатых химер (Rhinocnimaeridae), имеющих очень длинное заостренное рыло, также специфично дл€ материкового склона. Ќаиболее глубоководными среди 6 видов этой группы €вл€ютс€ гарриотты (род Harriotta), опускающиес€ до глубины 2600 м.
ѕерейдем теперь к костным рыбам. —овершенно особое место среди них занимает латимери€, или целакант (Latimeria chalumnae), Ч единственный современный представитель большой группы кистеперых рыб, когда-то Ч с нижнего девона (400 млн лет назад) до верхнего мела (65 млн лет назад) Ч процветавших в морских и пресных водоемах «емли, а затем полностью исчезнувших из палеонтологической летописи и считавшихс€ давным-давно вымершими. »менно поэтому поимка живого целаканта у берегов ёжной јфрики в 1938 г. стала одной из крупнейших биологических сенсаций нынешнего столети€, впрочем, эта истори€ слишком хорошо известна, чтобы еще раз пересказывать ее здесь (с ней можно ознакомитьс€, прочитав книгу проф. ƒж. —мита Ђ—тарипа Ч четвероногї).
—ейчас считаетс€ установленным, что латимерии, как это ни удивительно, нормально обитают только в одном и к тому же очень ограниченном участке ћирового океана Ч у бесшельфовых вулканических островов √ранд- омор и јнжуан (из группы оморских островов), расположенных в северной части ћозамбикского пролива, а перва€ из пойманных рыб лишь случайно заплыла так далеко на юг. насто€щему времени в руки ученых попало уже около сотни латимерии, и все они были пойманы коморскими рыбаками на традиционные примитивные удочки, предназначенные в основном дл€ лова руветты (жир этой рыбы используетс€ ими дл€ изготовлени€ противомал€рийной мази дл€ растирани€). ѕромысел ведут ночью с небольших лодок (типа каноэ с аутриггером), не удал€ющихс€ от берега больше чем на километр. Ётого достаточно, чтобы зан€ть место над свалом глубин и опустить крючок, наживленный куском прометеевой рыбы, в придонный слой. »менно там, среди скал на глубине от 70 до 600 м, лов€тс€ целаканты (местные жители называют их Ђгомбесса джомолеї), но чаще всего они берут приманку на глубине 150Ч300 м. ≈сть предположение, что ƒневные убежища латимерии приурочены к подводным пещерам, в которые по трещинам просачиваетс€ пресна€ вода с гор. ѕищу целакантов составл€ют рыбы (бентопелагические свет€щиес€ анчоусы, полимиксии и др.) и каракатицы. ќбычно добывают крупных латимерии длиной 90Ч200 см и массой 10Ч95 кг (предполагаетс€, что их максимальный возраст составл€ет около 10 лет), однако один –аз была добыта и совсем небольша€, по-видимому, годовала€ рыба, имевша€ всего 42 см в длину при массе 800 г, размножение происходит путем €йцеживорождени€ (как у многих акул) Ч в €ичнике одной самки были обнаружены 5 почти готовых к рождению эмбрионов длиной 30-33 см с еще не вт€нувшимис€ желточными мешками дна метром 8Ч13 см.
¬се остальные виды, вход€щие в состав океанических донно-придонных ихтиоцепов, относ€тс€ к костистым рыбам. –ассмотрим их в пор€дке системы, принимаемой сейчас большинством ихтиологов.
ќтр€д спиношипообразных полностью ограничен в своем распространении мезобатибенталыо. Ќотакантовые (сем. Notacanthidae) встречаютс€ во всех океанах в довольно широком диапазоне глубин (от 125 до 3700 м), но большинство видов т€готеет все-таки к верхней части склона (200Ч1300 м). ќни держатс€ в узком придонном слое (как показали подводные наблюдени€, эти рыбы плавают обычно в наклонном положении, так что голова находитс€ у самого грунта) и питаютс€ бентосными животными. Ќотаканты, или спиношипы (род Notacanthus) (рис. 34), особенно часто поедают мшанок, гидроидных полипов, губок, актиний, голотурий, а многошипы (род –о-lyacanthonotus) Ч ракообразных и полихет. »звестно, что у многих видов самки численно преобладают над самцами и имеют более крупные размеры, п это дает основани€ дл€ довольно правдоподобного предположени€ о возможном протандрическом гермафродитизме этих рыб. »х личинки Ч лептоцефалы имеют листовидно уплощенное удлиненное и прозрачное тело и достигают очень крупных размеров (до 1,8 м). ѕри метаморфозе длина, по-видимому, сильно уменьшаетс€, так как взрослые экземпл€ры большинства видов не превышают 50Ч60 см в длину (только Notacanthus chemnitzi может вырастать до 120 см).
—емейство галозавровых (Halosauridae), представители которого вылавливались на глубине 400Ч5000 м (обычно не глубже 3000 м, в абиссобенталь проникает только один вид рода Aldrovandia), мало отличаетс€ от нотакантовых по образу жизни. »х пищу также составл€ет бентос, но не седентарные (сид€чие) формы, а главным образом ракообразные и полихеты. ќдни виды предпочитают твердые грунты, другие Ч илистые участки. ќсоби некоторых видов, возможно, держатс€ стайками. —емейство насчитывает 16 видов, среди которых есть как циркумтропические формы (Halosauropsis macrochir, Aldrovandia affinis), более обычные в батибентали, так и узкораспространенные (многие виды рода Halosaurus), которые т€готеют к мезо-бентальным глубинам.
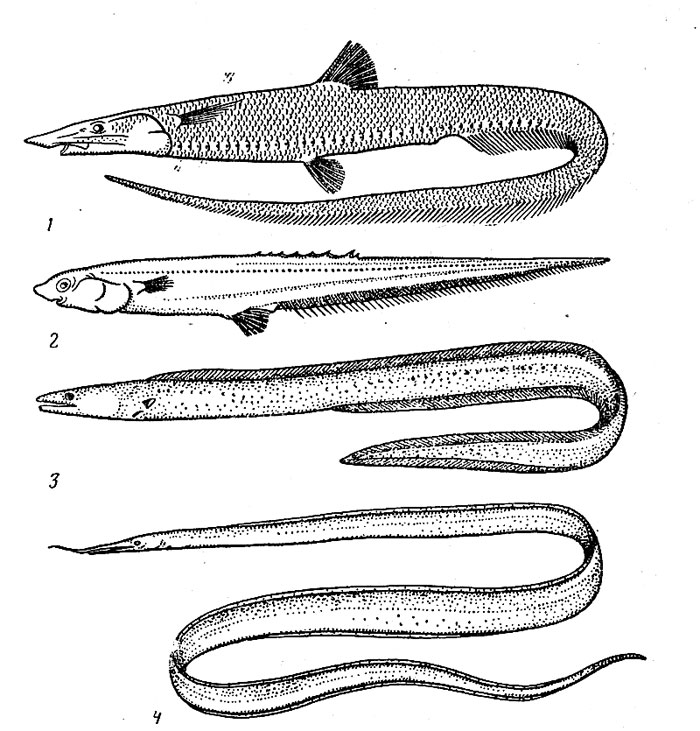
–ис. 34. јльдрованди€ (J), спиношип (2), слитножаберный угорь (3) и угорь венсфика (4).
ƒовольно обычны, а местами и многочисленны в батиальной зоне (преимущественно в пределах тропической области) донно-придонные угреобразные рыбы, принадлежащие к п€ти семействам (всего в отр€де их насчитываетс€ 16) и различающиес€ по глубине обитани€. “олько в верхней мезопелагиали Ч до глубины 300Ч400 м встречаютс€ отдельные представители преимущественно мелководных острохвостых угрей (сем. Ophichthidae), в том числе Ophisurus serpens Ч наиболее крупный вид семейства, достигающий 2,5 м в длину, а также ложномуреновых угрей (Xenocongridae). √лубже 500Ч600 м редко опускаютс€ и конгеровые угри (Congridae) Ч семейство, которое насчитывает около 30 родов и 100 видов, в большинстве своем шельфовых. Ќаиболее глубоководный из них Ч Coloconger scholesi, обитающий в западной части “ихого и в »ндийском океанах, живет, однако, па глубине 600-1200 м.
—пецифичны дл€ мезобатибентали всего два семейства угрей Ч неттастомовые (Nettastomatidae) и слитножабер-ные (Synaphobranchidae). первому из них принадлежат более 35 видов, из которых 10 известны сейчас только по личинкам Ч лептоцефалам. ¬зрослые рыбы достигают 1 м в длину. ¬се они имеют сильно выт€нутое рыло, у некоторых оканчивающеес€ м€систым щупальцевидным отростком. Ёти угри живут на глубине от 500 до 2000 м, причем чернохвоста€ неттастома (Nettastoma melanurum), а возможно, и другие виды устраивают норы в м€гком грунте. ќсновной их пищей €вл€ютс€ макропланктонные ракообразные.
—литножаберные угри обычно обитают в том же батиметрическом диапазоне, хот€ отдельные особи ловились и глубже 4000 м. —реди них есть как эпимезобентальные (виды рода Dissoma), так и чисто батибеитальные (Histiobranchus bathybius) формы; некоторые достигают крупных размеров (рекордна€ длина 1,8 м отмечена у Diastobranchus capensis из вод ёжной јфрики и јвстралии). Ёто Ч хищники и пожиратели падали, питающиес€ рыбой, головоногими моллюсками и ракообразными. ќдин из видов Ч обезь€ний угорь (Simenchelys parasiticus) нар€ду с потреблением мелких амфипод и копепод может, как предполагают, нападать на крупных рыб и, прогрыза€ стенку тела, выедать их внутренности (возможно, впрочем, что объектом Ђнападени€ї были в действительности уже погибшие рыбы). Ќекоторые из слитножаберных угрей встречаютс€ далеко за границами тропической зоны Ч вплоть до »сландии и Ѕерингова мор€, но эти районы могут, веро€тно, считатьс€ областью их стерильного выселени€.
»сключительно своеобразные одночелюстные угри (сем. Monognathidae), у которых в отличие от всех прочих рыб верхн€€ челюсть полностью редуцирована, так что формально их можно назвать челюстноротыми с большой нат€жкой, относ€тс€ тем не менее всего лишь к особому семейству, близкому к мешкоротам. ¬ ихтиологических коллекци€х разных стран сейчас насчитываетс€ около 75 экз. моногнатов, и все они были исследованы датскими учеными ». Ќильсеном и Ё. Ѕертельсеном, получившими исключительно интересные результаты. ѕочти все рыбы длиной от 4 до 15 см были пойманы в толще воды на глубине от 200 до 2000 м, но более часта€ встречаемость в придонных горизонтах позвол€ет предположительно относить ату группу к бентопелагическому ихтиоцепу, тем более что сама€ крупна€ особь длиной около 30 см была поймана у дна (этот экземпл€р, отличающийс€ от прочих черной окраской и хран€щийс€ ныне в «оологическом институте јЌ ———–, выловлен в районе урильских островов). —емейство включает 14 видов, многие из которых известны по единичным особ€м. ≈динственную пищу мопогнатов составл€ют крупные креветки, которых эти рыбы, вовсе не способные к быстрому плаванию и к тому же лишенные органов зрени€, обон€ни€ (обон€тельные розетки развиваютс€ только у половозрелых самцов) и сейсмосенсориой системы, конечно, не могут ловить активным путем. ѕредполагаетс€, что они подманивают добычу, выдел€€ в воду пахучий секрет специальной железы, расположенной па голове, и убивают близко подплывшую креветку, проткнув ее панцирь особым ростральным зубом, направленным вниз. Ётот непарный зуб имеет трубчатое строение (внутренний канал открываетс€ вблизи его острой вершины), и у его основани€ расположена железа, вырабатывающа€ €довитый секрет. ѕодобный аппарат, гомологичный зубам €довитых змей, €вл€етс€ совершенно уникальным среди рыб и рыбообразных.
ќтр€д лососеобразных представлен па глубинах несколькими группами. —емейство серебр€нковых (Argentinidae), содержащее около 35 видов, включает как бенто-пелагические, так и насто€щие пелагические формы, живущие на средних глубинах. водам, омывающим верхнюю часть материкового склона (на глубине 150Ч700 м, редко глубже), приурочены два рода Ч Argentina и Glossanodon. Ёто некрупные рыбы длиной 15Ч20 см, достигающие кое-где высокой численности. ѕочти все серебр€нки днем держатс€ у самого дна, а ночью ста€ми поднимаютс€ в толщу воды. Ћучше других изучена больша€ аргентина (A. silus), обитающа€ в умеренных водах —еверной јтлантики. ќна питаетс€ зоопланктоном (эуфаузииды, мелкие креветки и др.), имеет большую продолжительность жизни и достигает максимального размера (60 см) к 25 годам. ѕоловозрелой становитс€ в возрасте 10Ч12 лет, нереститс€ в апреле Ч июне, выметыва€ 3500Ч7500 крупных пелагических икринок. ¬торой род семейства Ч глоссанодоны, или малозубые серебр€нки, т€готеет к субтропическим водам.
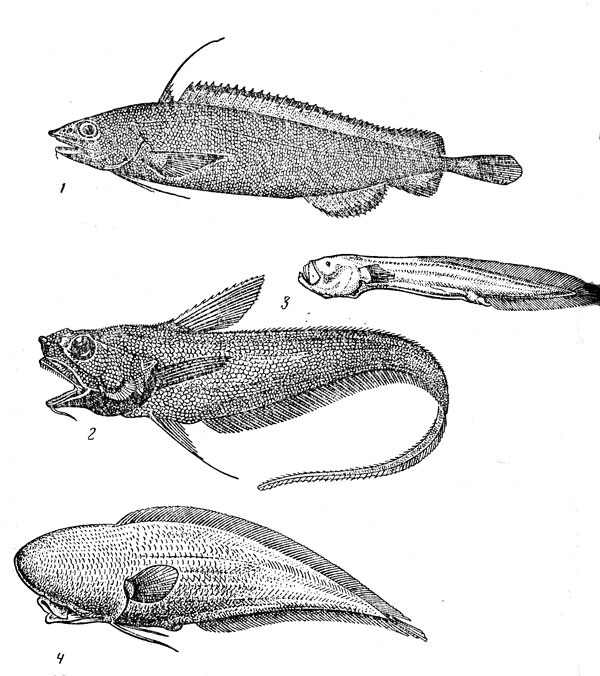
–ис. 35. √ладкоголововидные рыбы: гладкоголов (1), батиприон (2), фотостил (3), сирси€ (4).
√ладкоголововидные рыбы (рис. 35) Ч группа, относ€ща€с€ к числу самых характерных дл€ материкового склона и склонов подводных подн€тий. ¬ последние годы их изучение значительно продвинулось прежде всего благодар€ работам ё. ». —азонова и ƒ. ћаркла, но это относитс€ главным образом к систематике, так как сведений об экологии отдельных видов до сих пор недостает. семейству гладкоголовых (Alepocephalidae), не представленному только в јрктике и јнтарктике, принадлежит около 90 видов (наиболее разнообразна фауна тропической »ндо-¬естпацифики). ¬се они вход€т в состав бентопелагических ихтиоценов, однако их св€зь с дном выражена в разной степени. ќдни виды, составл€ющие преобладающее большинство (представители родов Alepocephalus, Conoсага и др.), приурочены к узкому придонному слою, другие, например Photostylus pycnopterus, обитают и в значительном удалении от склона, третьи (Microphotolepis) даже совершают суточные вертикальные миграции, поднима€сь ночью к границе эпипелагиали. ¬ целом диапазон батиметрического распространени€ семейства очень широк Ч от 100Ч150 до 5000 м, но наиболее обычны гладкоголовы на глубине 700Ч3000 м. ¬ семействе преобладают виды, т€готеющие к нижней батиали, а число верхнебатиальных значительно больше, чем абиссальных (к последним относ€тс€, по-видимому, роды Bellocia и Rinoctes), размеры гладкоголовов варьируют от 15Ч20 см до 1 м. ¬ их питании преобладают планктонные объекты (очень обычны, в частности, Ђстуденистыеї формы Ч медузы, гребневики, пиросомы, а также ракообразные), но нередко поедаетс€ и бентос (моллюски, полихеты, иглокожие). –ост медленный Ч южный гладкоголов (Alepocephalus australis), например, достигает длины 70 см в возрасте 20 лет. ѕлодовитость колеблетс€ от 20 икринок (у Rinoctes nasu-tus) до 10 тыс. икринок. ќни имеют очень большой диаметр (2Ч7 мм) и, по всей веро€тности, откладываютс€ на дно.
”никальной особенностью семейства платитроктовых (Platytroctidae), по внешнему виду очень сходных с гладкоголовами, служит наличие подкожного плечевого органа, продуцирующего свет€щуюс€ слизь. Ётот орган открываетс€ наружу маленькой кожной трубочкой, через которую слизь, содержаща€ люциферин и люциферазу, извергаетс€ наружу и распыл€етс€ в воде, где создает €ркое свет€щеес€ облачко голубовато-зеленого цвета. ” некоторых видов семейства есть и фотофоры, но не округлой формы, как у большинства других свет€щихс€ рыб, а в виде полосок-Ђшевроновї, пересекающих, как правило, нижнюю поверхность тела. ¬ семействе насчитываетс€ около 40 видов, из которых больша€ часть описана в последние 20Ч25 лет. ѕлатитроктовые Ч некрупные рыбы длиной от 12 до 30 см (наибольших размеров достигают представители рода Holtbyrnia). ¬ своем преобладающем большинстве они обитают в бентопелагиали на глубине 700Ч2000 м, и лишь один вид Ч Platytroctegen mirus, распространенный в северной части »ндийского океана, возможно относитс€ к числу мезопелагических. Ѕиологи€ этих рыб практически не изучена.
Ќемаловажна€ роль в бентопелагических ихтиоценах принадлежит и некоторым макропланктонным рыбам, вход€щим в состав таких семейств, в которых преобладают насто€щие пелагические виды, не имеющие никакой св€зи с дном. ¬ отр€де стомиеобразных такие виды есть среди гоностомовых (например, Triplophos hemingi и Gouostoma denndatum), топориковых (все 25 видов рода –оlyipnus, мавролик Maurolicus muelleri) (рис. 36), фотихто-вых (виды рода Polymetme), астронестовых (группа Astronesthes lucifer), меланостомиевых (два вида рода Odontostomias). ќсобое место в этом списке занимает мавролик (по всей веро€тности, под приведенным выше названием объедин€ют 5Ч6 близких видов), который €вл€етс€ одной из самых массовых рыб во многих участках материкового склона (в водах Ќорвегии он живет даже в глубоких фиордах) и у некоторых подводных подн€тий. Ќаибольша€ длина мавролика, широко распространенного над материковыми склонами и подводными подн€ти€ми во всех океанах, достигает 6Ч6,5 см. Ёто стайна€ рыба, держаща€с€ днем в придонных сло€х на глубине 200Ч600 м, но отрывающа€с€ от дна ночью и плотными кос€ками поднимающа€с€ в верхний 100-метровый слой. ћавролик созревает в годовалом возрасте и выметывает 200Ч500 довольно крупных икринок, причем размножение в умеренных водах обоих полушарий происходит в весенне-летнее врем€, а в тропиках идет круглогодично. ¬озраст этой рыбы не превышает 2Ч3 лет. ѕитаетс€ она копеподами и эвфаузиидами, а сама служит пищей многим хищникам.
–ис. 36. ћавролик (1), топорик полипнус (3) и морской бекас (3).
придонно-пелагическому комплексу средних глубин относ€тс€ и многие свет€щиес€ анчоусы. Ќаиболее известен среди них лампаниктод (Lampanyctodes hectoris), обитающий в водах материкового склона ёжной јфрики, јвстралии, Ќовой «еландии и „или и образующий в некоторых районах значительные скоплени€, которые используютс€ промыслом. ¬ темное врем€ суток эта небольша€ миктофида (длина до 6Ч7 см) совершает миграции в верхние слои воды и даже по€вл€етс€ у поверхности. јналогичный образ жизни ведет Diaphus suborbitalis (наибольша€ длина 8 см), распространенный у берегов ¬осточной јфрики, »ндонезии и японии (он особенно многочислен в заливах —агами и —уруга), а также над многими подводными подн€ти€ми в »ндийском и “ихом океанах. Ќужно отметить, что во врем€ ночного подъема диафус удал€етс€ на 200Ч300 м от дна, иногда пересека€ разнонаправленные течени€, и остаетс€ тем не менее над своим подн€тием, ко дну которого он вновь опускаетс€ утром. ак видно, эта рыба, во-первых, может ориентироватьс€ в пространстве, не име€ зрительного контакта с неподвижным субстратом, и, во-вторых, способна противосто€ть течени€м, имеющим скорость пор€дка 1Ч2 узлов (т. е. 1,8Ч3,6 км/ч), чтобы сохран€ть свое положение относительно горы. —пособ ориентации остаетс€ пока неизвестным. ¬о всех остальных отношени€х этот вид не отличаетс€ от мигрирующих мезопелагических миктофид: он питаетс€ зоопланктоном и имеет короткий жизненный цикл Ч созревает к концу первого года жизни, а затем нереститс€, выметыва€ порцию за порцией икру в течение нескольких мес€цев вплоть до естественной гибели. »з других бентопелагических свет€щихс€ анчоусов, совершающих суточные миграции, следует упом€нуть такие виды, как D. taaningi и Benthosema fibulatum, достигающие высокой численности (первый Ч над материковым склоном обоих берегов јтлантики, второй Ч в »ндийском и “ихом океанах). —реди миктофид есть и виды, посто€нно приуроченные к очень узкому придонному слою и не покидающие его даже ночью. Ёти виды, например Diaphus adenomus и D. coeruleus, лов€тс€ только донными тралами.
ѕридонные виды известны также в таких преимущественно пелагических группах, как семейство меламфаевых (Melamphaes acanthomus обитает только над склоном западного побережь€ јмерики от алифорнии до 20∞ ю. ш.) и подотр€д глубоководных удильщиков, в котором все 4 вида семейства дицерациевых (Diceratiidae) приурочены к мезобентопелагиали, а представители рода Thaumatichthys (сем. Thaumatichthyidae) вообще ведут донный образ жизни. “ауматихт (“. axeli), известный с глубины до 3600 м, поджидает свою добычу, которую он привлекает свет€щейс€ приманкой, расположенно внутри ротовой полости, лежа на дне, и заглатывает ей не предпринима€ практически никаких усилий.

–ис. 37. √лубоководные аулопообразныс рыбы: ипнопс (1), батитифлопс (2) батиптер (3), бентозавр (4).
ќтр€д аулопообразных представлен на всех глубинах бентального биотопа. ¬ верхней части склона обычны зеленоглазки (род Chlorophthalmus), обитающие в широко батиметрическом диапазоне (от 100Ч150 до 1000 м). Ёти небольшие (длина до 20 см) рыбы, которые ведут станую жизнь вблизи грунта и питаютс€ бентосом. ак показано . —удаком, к этому же семейству (Chlorophtha midae) принадлежат несколько родов, очень характерных дл€ больших глубин (рис. 37). Ѕатиптеры (около 20 видов рода Bathypterois длиной 15Ч30 см) встречаютс€ в всех океанах на глубине 250Ч6000 м. ѕодводными наблюдени€ми и фотосъемкой установлено, что они обычно неподвижно сто€т на дне, возвыша€сь над ним, как на ходул€х, на утолщенных и удлиненных краевых лучах брюшных плавников и хвоста (их называют еще рыбами-треногами), и выжидают по€влени€ планктонных рачков Ч их основной пищи. ƒлинные лучи грудных плавников выполн€ют при этом роль чувствительных антенн, так как глаза у этих рыб развиты плохо. ¬ отличие от разнополых зеленоглазок батиптеры €вл€ютс€ синхронными гермафродитами, как и другие глубоководные виды семейства Ч батимикропсы (Bathymicrops), батитифлопсы (Bathytyphlops) и ипнопсы (Ipnops), обитающие в батин абиссопелагиали. ” представителей последнего рода глаза полностью редуцированы, а роль светочувствительных органов выполн€ют так называемые глазные пластины, расположенные на верхней поверхности головы под тонкими кост€ми. ѕлодовитость батитифлопсов достигает, по Ќ. ћерритту, 6000 икринок, проход€щих развитие в толще воды: их личинки, описанные как особый род Macristiella, живут в эпипелагиали. »ныопсы и батимикропсы откладывают не более 300 икринок, которые имеют отрицательную плавучесть. Ёти рыбы, по-видимому, провод€т у дна всю свою жизнь.
¬ близком семействе €щероголововых (Synodonlidae) больша€ часть видов живет на шельфе, хот€ некоторые зауриды (род Saurida) встречаютс€ и в верхней мезобентали Ч до глубины 550 м. Ћишь два вида рода Bathysaurus относ€тс€ к числу насто€щих глубоководных рыб Ч один из них (¬. mollis) ловитс€ преимущественно па глубине 2500Ч4500 м при температуре 2Ч3∞. Ѕатизавры Ч крупные хищные рыбы (длина до 85 см), питающиес€ в основном рыбой. ќни тоже синхронные гермафродиты, плодовитость составл€ет 24Ч56 тыс. икринок. Ќа личиночно-мальковой стадии развити€ (ранее также относившейс€ к особому роду Macristium) держатс€, по-видимому, в мезопелагиали. ќсобое место среди €щероголовов занимают придонно-пелагические бомбили Ч род Harpodon, из трех видов которого наиболее известна Ђбомбейска€ уткаї (Ќ. nehereus) длиной до 30 см, широко распространенна€ в прибрежных водах »ндийского и западной части “ихого океанов. ¬ определенные сезоны эта рыба ста€ми по€вл€етс€ вблизи берегов и даже входит в опресненные эстуарий, где ее вылавливают в немалом количестве, но значительную часть жизни она, как предполагают, проводит на глубинах склона.
ѕо внешнему облику бомбейска€ утка вполне соответствует представлению о глубоководной рыбе, и тем более неожиданной дл€ мен€ была перва€ (и единственна€) встреча с ней... ¬ 1961 г Ђ¬ит€зьї уходил из алькутты, крупнейшего порта »ндии, расположенного на одном из рукавов дельты √анга Ч реке ’угли, воду которой можно сравнить только с глинистым раствором: она им€ ет цвет кофе с молоком и, подобно нашим среднеазиатским рекам несет огромное количество взвеси. Ќа выходе в такой же мутный эстуарий к борту подошла небольша€ парусна€ лодка с двум€ рыбаками-бенгальцами, предложившими свой скромный улов несколько рыб, аккуратно уложенных в плетеную корзину. ¬ этом то улове (мы получили его в обмен на сгущенное молоко) оказались бомбили. Ѕесчешуйные большеротые Ђуткиї отличались др€блой консистенцией жирного тела, очень характерной дл€ многих глубоководных рыб, и буро-коричневой окраской. Ёти. Ђптицыї, надо сказать пр€мо, не очень-то наход€т место в той экологической классификации, котора€ прин€та в этой книге - ведь из числа океанических рыб в эстуари€х можно встретить, кроме них, пожалуй, только брегмацеров (один из них, кстати, был даже описан по экземпл€ру из эстуари€ ’угли).
«аверша€ обзор аулопообразных, надо отметить, что к числу бенто-пелагических рыб нужно относить и преобладающее большинство скопелозавров (род Scopelosaurus из сем. Notosudidae), взрослые особи которых держатс€ только над континентальными склонами, островными цокол€ми и подводными подн€ти€ми.
ќчень много донно-придонных океанических видов содержит отр€д трескообразных рыб, две трети состава которого обитает на значительных глубинах. ѕреимуществ венно глубоководным €вл€етс€, в частности, семейство моровых (Moridae), впервые обособленное известным отечественным ихтиологом ј. Ќ. —ветовидовым (1903Ч1985).
—удьба довольно близко свела мен€ с јнатолием Ќиколаевичем —ветовидовым. я познакомилс€ с ним еще в середине 50-х годов, при одной из первых командировок в ленинградский «оологическпй институт. Ёту встречу помню хорошо. ј. Ќ. пригласил мен€ в кабинет и, вскользь вы€снив, кто €, откуда и чем интересуюсь, целиком вз€л нить разговора в свои руки и долго с большим увлечением говорил о своих студенческих годах (он училс€ на ќтделении рыбоведени€ ћосковской сельскохоз€йственной академии, из которого впоследствии образовалс€ ћосрыбвтуз, так что, получалось, мы окончили один вуз), о своих старших коллегах учител€х Ч ¬. . —олдатове, Ќ. ћ. ниповиче и Ћ. —. Ѕерге, которых глубоко чтил, о детал€х своей тогдашней работы по черноморским рыбам. ѕри последующих посещени€х Ћенинграда эти беседы-монологи вошли в обычай (видимо, € оказалс€ хорошим слушателем), а через несколько лет € стал вхож и в квартиру на ћосковском проспекте (у ј. Ќ. была прекрасна€ ихтиологическа€ библиотека) и на дачу в омарове, познакомилс€ с добрейшей ≈лизаветой ¬асильевной, стал иной раз останавливатьс€ у —ветовидовых при коротких приездах (ј. Ќ. всегда выговаривал мне, если € находил иное пристанище). — середины 70-х годов наши встречи стали очень регул€рными: будучи членом бюро ќтделени€ общей биологии јкадемии наук, ј. Ќ. почти еженедельно бывал в ћоскве и постепенно у него вошло в привычку проводить у мен€ дома последние четыре-п€ть часов в ожидании полуночного курьерского поезда. ¬се это врем€ он рассказывал, рассказывал и рассказывал...
ћен€ всегда поражали трудолюбие и организованность ј. Ќ., который как бы сам задавал себе ежедневный урок и всегда выполн€л его независимо ни от чего. “ри капитальные монографии Ч Ђ—ельдевыеї, Ђ“рескообразныеї и Ђ–ыбы „ерного мор€ї, вошедшие в золотой фонд ихтиологии,Ч итог упорного многолетнего труда с полной отдачей сил и времени. ќгромна€ требовательность к себе позвол€ла ј. Ќ. быть чрезвычайно требовательным и к своим сотрудникам и ученикам, и он не прощал никому из них ни отступлений от генеральной линии исследований, ни отвлечений на посторонние, с его точки зрени€, дела, ни тем более про€влений неусидчивости или недисциплинированности. ≈го отношени€ с коллегами по институту не были легкими, а с некоторыми из них он, оставив административную де€тельность, не общалс€ вовсе. ѕо-видимому, именно поэтому он искал и находил себе собеседников на стороне и открывалс€ им с самой неожиданной стороны Ч ведь его монологи касались не только профессиональных вопросов, но также истории, литературы, искусства. ¬прочем, с любой темы ј. Ќ. с неизбежностью возвращалс€ к ихтиологии, котора€, несомнено, была единственной насто€щей страстью его жизни.
ћоровые рыбы (их около 90 видов) близкородственны тресковым, но обитают в основном на глубинах материкового склона (в семействе есть, впрочем, и шельфовые виды, распространенные преимущественно в умеренных широтах южного полушари€, а представители 3Ч4 монотипических родов живут, как предполагают, в толще воды). ќни представлены во всех океанах и особенно многочисленны в верхней батиали, но некоторые виды встречаютс€ в более широком диапазоне глубин Ч антимора (Antimora rostrata) (рис. 38), например, от 400 до 3000 м. ƒлина океанических морид варьирует от 15Ч25 см у физикулюсов (род Physiculus) до 80-90 см у лепидионои (Lepidion). ќни сильно различаютс€ по образу жизни.‘изикулюсы, подобно налимам, ищут свою добычу на дне, пользу€сь обон€нием (поэтому они часто попадают в донные ловушки); антимора поедает как бентосных животных, которых она может выкапывать из м€гкого грунта лопатообразным рылом, так и придонно-пелагических рыб и кальмаров; дальневосточна€ лемонема (Laemonema longipes) совершает суточные вертикальные миграции, поднима€сь ночью в пелагиаль, и питаетс€ там; макропланктоном, крупные лепидионы и моры (род Morа) ед€т креветок, рыб и кальмаров.
–ис. 38. √лубоководные трескообразные и ошибнеобразные рыбы: антимора (1) долгохвост (2), нибеллинелла (3), слепой тифлонус (4)
Ѕиологи€ глубоководных морид в целом изучена недостаточно. »сключение составл€ет, пожалуй, только лемонема, обитающа€ в северной части “ихого океана от берегов ’онсю до залива јл€ска на глубине 400Ч1200 м. Ћемонема Ч тугоросла€ рыба, созревающа€ при длине более 40 см в возрасте 5Ч8 лет и достигающа€ предельной длины около 70 см в возрасте 26 лет. Ќачальные годы жизни она проводит в толще воды, а затем опускаетс€ в придонные горизонты (по ночам мигриру€ в пелагиаль), причем более крупные особи держатс€ в нижней части батиметрического диапазона, занимаемого видом. ƒальневосточной лемонеме свойственны значительные сезонные перемещени€: летом она концентрируетс€ в основном на склонах восточного ’оккайдо и урильских островов, а зимой уходит в воды ёжной японии, где нереститс€, выметыва€ 300Ч800 тыс. мелких икринок, на глубине 600Ч800 м. Ёта рыба служит объектом промысла.
ƒл€ самой верхней части склона, пограничной с шельфом, характерны также мерлузы, или хеки (род Merluccius), и макруронусы (Macruronus) Ч хищные рыбы, достигающие 0,7Ч1,3 м (все они имеют промысловое значение), из которых лишь немногие могут быть встречены вплоть до глубины 1000 м. ƒругие представители семейства Merlucciidae, например штайндахнери€ (Steindaclmeria argentea) из западной тропической јтлантики, имеюща€ сложную систему свет€щихс€ органов, €вл€ютс€ па-сто€щими мезобентопелагическими рыбами.
—ледующее семейство отр€да трескообразных Ч длиннохвостовые, или макруриды (Macrouridae), Ч сама€ многообразна€ группа глубоководных рыб, насчитывающа€ более 300 видов. ¬ последние годы она интенсивно изучалась ¬. ћ. ћакушном, Ќ. ћаршаллом, “. »вамото, ё. ». —азоновым, ƒ. —тейном, ќ. ќкамура и другими исследовател€ми. ћакруриды широко распространены в ћировом океане (они отсутствуют только в јрктике) на глубине от 50Ч100 до 5Ч6 тыс. м. „исто шельфовых видов в семействе, впрочем, очень немного, и все они принадлежат к роду полорылов Ч Coelorinchus). ≈стественно, отдельные виды имеют более узкое географическое и батиметрическое распространение. ѕреобладающее большинство долгохвостов приурочено к мезобентали и мезобентопелагиали (200Ч1500 м), причем многие из них в высшей степени стенобатны: у р€да видов незумий (род Nezumia) и гименоцефалов (Hymenocephalus) весь диапазон вертикального распределени€ укладываетс€ в две-три сотни метров. ƒл€ нижней батиали (т. е. батибентали) особенно характерны роды Macrourus и Coryphaenoides (см рис. 38), а в абиссальную зону проникают лишь отдельные представители последнего (например, —. armatus), а также роды Echinomacrurus и Astheriomacrurus. ƒл€ верхнебатиальных видов очень типичен значительный региональный эндемизм (иначе говор€, они имеют ограниченное географическое распространение), но в более глубоких горизонтах ареалы долгохвостов, как правило, занимают обширные площади. Ѕольшинство видов обитает в тропической зоне.
¬нешний вид долгохвостов чрезвычайно своеобразен: у них непропорционально больша€ голова и очень длинный постепенно утончающийс€ в нить хвост, за который англичане называют этих рыб Ђкрысохвостамиї. ѕодобно отдельным моровым и мерлузовым рыбам, некоторые роды макрурид обладают одиночными или, реже, парными свет€щимис€ железами, расположенными на брюхе перед анальным отверстием и содержащими бактерий-симбионтов, излучающих свет в результате своей жизнеде€тельности. “ака€ железа состоит из мешковидного резервуара дл€ бактерий, светоотражающего сло€ и прозрачной линзы. ќна снабжаетс€ кровью через систему капилл€рных сосудов, обеспечивающих бактерии кислородом, и светоиспускание может регулироватьс€ как изменением интенсивности кровеснабжени€, так и Ђзашториваниемї линзы темными хроматофорами. Ќекоторые виды (например, м€гкоголовый долгохвост Malacocephalus laevis), по-видимому, способны выпускать в воду струю свет€щейс€ жидкости.
ƒолгохвосты сильно различаютс€ по размерам. —амые мелкие из них не превышают 15Ч20 см, а самые крупные достигают 2 м в длину. —реди них есть как насто€щие донные формы, в поисках пищи перекапывающие грунт жесткими носами (жесткорылы рода Trachyrinchus и многие полорылы), так и питающиес€ макропланктоном бентопелагические формы, составл€ющие €вное большинство, а также немногочисленные чисто пелагические виды, которые упоминались в предыдущей главе. ¬се эти категории св€заны переходами, так как многие долгохвосты могут быть встречены как на грунте, так и в отрыве от него. Ќекоторые виды из рода Coryphaenoides (тупорылый макрурус —. rupestris и другие) даже совершают регул€рные миграции с подъемом в пелагиаль в ночно€ врем€, но никогда не поднимаютс€ в верхние слои воды
Ѕолее или менее полные представлени€ об экологии существуют сейчас только в отношении тех долгохвостов, которые уже вовлечены в промысловое использование. их числу относ€тс€, в частности, северотихоокеапские малоглазый (Coryphaenoides pectoralis) и пепельный (—. cinereus) макрурусы, а также североатлантический тупорылый макрурус. ¬се они обитают в широком диапазоне глубин (от 140Ч180 до 2,5Ч3,5 тыс. м), но в массовом количестве встречаютс€ только в верхней части материкового склона. Ќаибольша€ длина этих видов 0,5Ч2,0 м, предельный возраст 10Ч40 лет. ¬ соответствии с размерами находитс€ и плодовитость: малоглазый макрурус откладывает до 400 тыс. икринок, тупорылый Ч до 60 тыс., пепельный Ч до 18 тыс. Ќесмотр€ на довольно высокую численность этих видов, их планктонные личинки попадаютс€ в пробах очень редко, и местообитание ранних стадий развити€ остаетс€ неизвестным. ќсновную пищу всех трех видов составл€ет мезопелагический макропланктон Ч креветки, криль, кальмары, а из рыб Ч свет€щиес€ анчоусы, значительно меньша€ роль принадлежит бентосу, хот€ малоглазый макрурус, например, почти не выходит в пелагиаль.
’орошо представлено в глубинных бентопелагических комплексах семейство ошибневых (Ophidiidae), по видовому разнообразию уступающее одним только долгохво-стам. —юда относитс€ около 200 видов (достойный вклад в их изучение внесли ». Ќильсен, ƒ. оэн, ё. Ќ. ўер-бачев), и почти три четверти из них ведут глубоководный образ жизни. —реди ошибневых есть верхнебатиальные формы, в частности мономитопы (род Monomitopus) и многие дикролены (Dicrolene), но особенно характерны они дл€ нижней части склона и дл€ ложа океана, где живет большинство видов семейства, в том числе наиболее мелкие (Tauredophidium hextii длиной немногим более 10 см) и наиболее крупные (Spectrunculus grandis длиной 1,3 м и массой 16,5 кг) его представители. числу самых глубоководных видов принадлежат слепой тифлопус (Typhlonus nasus) (см. рис. 38), обнаруженный вплоть ƒо глубины 5100 м, лентовидный бассоцет (Bassozetus taenia) Ч до 5600 м, атлантический лейцикорус (Leucicorus atlanticus) Ч до 6800 м, холкомиктеронус (Holcomycteronus profundissimus) Ч до 7160 м и абиссобротула (Abyssobrotula galatheae), которой принадлежит абсолютный рекорд по глубине вылова: в желобе ѕуэрто-–ико она была добыта с глубины 8370 м. ¬се глубоководные офидииды во взрослом состо€нии лов€тс€ только донными тралами, но их личинки, по-видимому, живут в толще воды. ѕищу этих рыб, нигде не достигающих особенно большой численности, составл€ют донные животные, в основном полихеты и ракообразные.
—ходный с офидиидами образ жизни ведут представители близкого к ним семейства бититовых (Bythitidae), которые отличаютс€ способностью к живорождению (оплодотворение у них, естественно, внутреннее, и самцы имеют довольно сложно устроенный копул€тивпый орган). ¬иды катетиксов (род Cataetyx), в частности, довольно обычны на всех глубинах континентального склона и встречаютс€ во всех океанах. ¬зрослые особи этих видов живут на самом дне, а молодь, заметно отличающа€с€ от взрослых по внешнему виду, держитс€ в толще воды. Ѕентопелагические афионовые рыбы (сем. Aphyonidae) длиной от 4 до 18 см Ч обитатели придонных горизонтов, встречающиес€ в нижней батиали и над ложем океана (наибольша€ глубина поимки Ч 5320 м ).Ќекоторые нетопыревые рыбы встречаютс€ и гораздо глубже.
—ледующий отр€д Ч удильщикообразные Ч характерен преимущественно дл€ мезобентали, в которой он представлен трем€ семействами и примерно сотней видов. Ѕольшинство удильщиков (сем. Lophiidae), которых называют также морскими черт€ми, обитает на глубине до 500Ч600 м, но некоторые, например европейский морской черт (Lophius piscatorius), могут опускатьс€ гораздо глубже - даже до 2000 м. Ёти своеобразные рыбы длиной 0,3Ч1,2 мм имеют уплощенное тело, непропорционально большую голову и огромную зубастую пасть и провод€т большую часть жизни, лежа на дне в пассивном ожидании добычи (в основном различных рыб), которую подманивают с помощью особой вымпеловидной приманки Ч эски, вывешенной перед ртом на конце Ђудилищаї Ч длинного обособленного луча спинного плавника. ¬ том же диапазоне глубин (200Ч500 м) обычны другие Ђуд€щиеї рыбы, также относ€щиес€ к числу бентических Ч жабовидные хаунаксы (сем. Chaunacinae) длиной до 40 см и морские нетопыри (Ogcocephalidae) (“ак, голотип Halieutopsis andriashcvi Ч вида, недавно описанного из тропической зоны »ндийского океана, был выловлен в батибентали на глубине около 4000 м.), редко превышающие 15Ч20 см в длину. ѕоследние во взрослом состо€нии практически тер€ют способность к плаванию: они перемещаютс€ по дну только по-пластунски Ч ползком, использу€ дл€ этого свои Ђрукиї Ч грудные плавники, как бы согнутые в локт€х, и Ђногиї Ч брюшные плавники и лишь подталкива€сь небольшим хвостовым плавником. ¬се эти рыбы, по-видимому, откладывают икру на дно, но их личиночное развитие идет в толще воды. ¬еро€тно, они имеют большую продолжительность жизни (один из некрупных видов морских нетопырей, к примеру, живет не менее 9 лет).
—реди преимущественно пелагических видов отр€да опахообразных особое место занимают донные ателеоповые рыбы (сем. Ateleopodidae), внешне очень сходные с долгохвостами (их истинное место в системе было установлено в самое недавнее врем€). Ќекоторые из них (представители рода Ijimaia) достигают крупных размеров (длина до 2 м) и широко распространены в тропической зоне на глубине от 200 до 1000Ч1500 м, хот€ повсюду относ€тс€ к числу редких рыб.
Ђ нижноеї Ч по описани€м и рисункам Ч знание рыб, конечно, позвол€ет узнавать этих рыб при встрече, но их истинный облик может очень сильно отличатьс€ от того представлени€, которое сложилось до личного знакомства. ƒолжен сказать еще, что мы, ихтиологи, обычно различаем уже известные нам виды не по их официальным диагностическим признакам, а по общему облику (как говоритс€, габитуально), т. е. так, как это делают все рыбаки (многие из них, пожалуй, не смогут ответить на вопрос, чем отличаетс€ лещ от густеры, но опознают их вполне уверенно).
— ателеопом мне довелось впервые встретитьс€ совсем недавно - только в мае 1987 г., когда Ђѕрофессор Ўтокманї проводил исследовани€ на подводном хребте —ала-и-√омес. ≈стественно, € вполне определенно представл€л себе эту уникальную рыбу, но действительность превзошла все ожидани€. ”видев в разв€занном куте трала среди €рких колючих антигоний, тригл и окуней белесый свинцового цвета глаз с крохотным темным окошечком в центре, € сразу обратил на него внимание. √лаз был окружен чем-то гладким и студнеобразным, и € только успел подумать об осьминоге, как ёрий —азонов поставил диагноз: Ђјтелеопус попалс€!ї (он уже видел этих рыб, работа€ на траулере в »ндийском океане). “ри наших атолеопа, пойманные па глубине 380 м, имели чуть больше полуметра в длину. ѕокрытые гладкой скользкой кожей, они больше всего- напоминали огромных головастиков. Ќаибольшее впечатление производила их голова Ч совершенно не рыбь€, м€гка€ и полупрозрачна€ как желе, обт€нута€ нежной скользкой кожей. ћаленький, воронкообразный и совсем беззубый ротик, расположенный на нижней поверхности, вызывал сильное сомнение в способности его обладател€ питатьс€ Ђрыбой, офиурами и ракообразнымиї (именно эти объекты находили в желудках ателеопид). ѕо необычности облика ателеопы нисколько не уступают ни удильщикам, ни хаулиодам, ни мешкоротам.

–ис. 39. √лубоководные бериксообразвые рыбы: стефаноберикс (1), средиземноморский хоплостет (2), диретмоид (3), низкотелый берикс (4).
—ущественна€ роль в ихтиоценах материкового склона и подводных подн€тий принадлежит некоторым представител€м отр€да бериксообразных рыб (рис. 39). Ќаиболыним разнообразием среди них выдел€етс€ семейство трахихтовых (Trachichthyidae), детально изученное в последние годы ј. Ќ. отл€ром. ѕочти все представители этой группы (а их около 40) относ€тс€ к числу полуглубоководных рыб, весьма обычных в нижней части шельфа и над его свалом на глубине 150Ч800 м. ќдин из наиболее широко распространенных видов Ч атлантический большеголов, или пилобрюх (Hoplostethus atlanticus), встречаетс€ у обоих берегов јтлантического океана и в умеренно теплых водах южного полушари€ на глубине от 300 до 1500 м (чаще всего 800Ч1200 м). Ётот вид, как и другие трахихтовые, характеризуетс€ медленным ростом (годовые приросты пор€дка 2Ч3 см) и большой продолжительностью жизни Ч до 24 лет. ѕоловой зрелости он достигает в возрасте 10Ч15 лет (плодовитость варьирует от 20 до 200 тыс. икринок), а наиболее крупные особи имеют длину около 60 см при массе около 4,5. ≈го пищу составл€ют макропланктонные животные: ракообразные, мелкие кальмары и рыбы.
¬ придонных сло€х воды над материковым склоном и подводными подн€ти€ми (обычна€ глубина 200Ч800 м) обитают бериксовые рыбы (сем. Berycidae). Ќизкотелый берикс (Beryx splendens) Ч €рко-красна€ стайна€ рыба, внешне напоминающа€ морского окун€ и достигающа€ 45 см в длину, встречаетс€ во всех океанах. ¬ дневное врем€ он держитс€ над дном, а ночью отрываетс€ от него и выходит в водную толщу, нередко образу€ значительные скоплени€. ѕищу берикса составл€ет различный макропланктон, преимущественно ракообразные, рыбы (в том числе миктофиды), мелкие кальмары.
»з других придонных бериксообразных нужно отметить диретмоидов (род Diretmoides), которые вместе с мезопелагическим диретмом (см. гл. III) вход€т в состав семейства Diretmidae. ќни достигают 40Ч50 см в длину, окрашены в темные тона и во взрослом состо€нии посто€нно пребывают над дном на глубине пор€дка 500Ч 800 м, хот€ их серебристые мальки, очень похожие на диретма, ведут пелагический образ жизни. ¬ нижней батиали живут также гиспидоберикс (Hispidoberyx ambagiosus), совсем недавно (в 1980 г.) описанный ј. Ќ. отл€ром и выделенный им в особое семейство, и гибберихты Ч два вида рода Gibberichthys. Ћичинки последних при длине 1Ч2 см резко отличаютс€ по внешнему виду от взрослых рыб и до последнего времени считались принадлежащими к самосто€тельному семейству. ” этих рыбок сильно удлиненный третий луч брюшных плавников имеет вид длинного стержн€, несущего около 30 больших листовидных мешков черного цвета с прозрачным овальным окошечком па конце каждого. Ёти придатки внешне очень напоминают некоторых сифонофор или медуз с их плавательными пузыр€ми и свисающими щупальцами и могут расцениватьс€ как про€вление мимикрии (не исключено, впрочем, что они испускают свет и служат дл€ приманивани€ добычи). Ћичинки и мальки гибберихтов держатс€ в поверхностных сло€х воды, взрослые рыбы живут в придонных горизонтах на глубине 750Ч2000 м.
—реднеглубинные бентопелагические рыбы есть и в отр€де солнечникообразных. ќдни из них Ч антигонии (род Antigonia) и солнечники (Zeus и Zenopsis) имеют центром батиметрического распределени€ зону, переходную между эпи- и мезопелагиалью (глубины пор€дка 100Ч400 м), другие, принадлежащие к семействам ореосомовых (Oreosomatidae) и чешуеиглых (Grammicolepididae), встречаютс€ только в верхней батиали Ч на глубине от 200Ч500 до 1500Ч2000 м. јллоцит, или лунник (Allocyttus verrucosus), длиной до 38 см широко распространен в јтлантике и южной части »ндийского океана и кое-где (например, на банке јгуль€с) даже образует плотные концентрации. ѕитаетс€ эта рыба разнообразным макропланктоном.
Ѕенто-пелагический образ жизни в диапазоне глубин от 100Ч200 до 600 м ведут морские бекасы (сем. ћасrоrhamphosidae), принадлежащие к отр€ду колюшкообразных. Ќаиболее обычный вид Ч морской бекас (ћасrоrhamphosus scolopax) длиной до 19 см Ч широко распространен в тропических и субтропических водах и передко встречаетс€ в огромном количестве и очень плотными ста€ми над относительно мелководными подводными подн€ти€ми. —читаетс€, что молодые бекасы, живущие в отрыве от дна, имеют более прогонистое тело и окрашены с преобладанием синих тонов, тогда как у взрослых тело выше, а в окраске больше красного цвета, но эта точка зрени€ не общеприн€та (“. ларк считает, что существуют два вида этих рыб).
√лубоководные виды имеютс€ в р€де семейств скорпенообразных рыб, более характерных дл€ прибрежной зоны. ¬ своем преобладающем большинстве они приурочены к самым верхним горизонтам мезобентали, пограничным с эпибенталью. ¬плоть до глубины 300Ч500 м (иногда и более) вполне обычны, в частности, некоторые скорпеновые рыбы (сем. Scorpaenidae) Ч морские окуни (Sebastes), беспузырники (Helicoienus) и др. ¬се они во взрослом состо€нии ведут бентический образ жизни (молодь встречаетс€, однако, в толще воды) и редко отрываютс€ от дна, но из этого правила есть исключени€. —уточные вертикальные миграции с ночным подъемом к поверхности отмечены у многих видов, а североатлантический окунь-клювач (S. mentella) вполне способен существовать в пелагиали, образу€ значительные скоплени€ даже над океаническими глубинами. »з тропических видов наиболее Ђпелагизированї батиально-пелагический Ectreposebastes imus, имеющий в отличие от других среднеглубинных скорпенид черную, а не красную окраску. насто€щим мезобеитальным рыбам принадлежат, однако, немногие скорпеновые, посто€нно живущие на глубине пор€дка 1000Ч1600 м (Ђрекордна€ї глубина 2500 м) Ч шипощеки (род Sebastolobus), обитающие в северной части “ихого океана, и трахискорпии (Trachyscorpia). –азмеры глубоководных скорпенид от 15Ч20 см до 1 м (обычно не крупнее 50 см). ¬се они, по-видимому, относ€тс€ к числу медленнорастущих, долгоживущих (до 10Ч 25 лет) и поздно созревающих рыб с невысоким темпом воспроизводства.
ƒл€ вод материкового склона характерно также семейство аноплопомовых (Anoplopomatidae). ”гольна€ рыба (Aiioplopoma fimbria), достигающа€ длины 1,2 и массы 14 кг, обитает в северо-восточной ѕацифике (от алифорнии до јл€ски), где встречаетс€ на всех этапах жизненного цикла. ≈сть она и в Ѕеринговом море (там, по-видимому, существует зависима€ попул€ци€ вида), а отдельные особи изредка лов€тс€ даже у урильских островов и японии. ”гольна€ рыба обитает на глубине 200Ч1200 м (максимальна€ глубина вылова 1800 м) и адаптирована к характерной дл€ этих глубин пониженной концентрации растворенного кислорода. Ћичинки и мальки держатс€ в эпипелагиали, годовики длиной 20Ч40 см переход€т к жизни у дна. √лавную пищу составл€ют рыба, головоногие моллюски и кишечнополостные (гребневики и медузы). ¬торой вид семейства Ч эрилепис (Erilepis zonifer), также обитающий в северной части “ихого океана, принадлежит, веро€тно, к верхне-мезобенто-пелагическому ихтиоцену.
Ќесколько мезобентальных видов содержит холодноводное семейство рогатковых (Cottidae), н все они имеют некрупные размеры (не более 15Ч20 см). “ак, обычные на севере “ихого и јтлантического океанов и в мор€х јрктики ицелы (род Icelus) и триглопсы (Triglops) встречаютс€ у дна вплоть до глубины 500Ч900 м, а самый глубоководный представитель группы Ч дальневосточный крючкорог Zesticelus profundorum Ч ловилс€ даже на глубине свыше 1000 м. Ќа материковом склоне живут и австрало-новозеландские рогатки-антиподы (род Antipodocottus), а также единственный экваториальный вид группы Ч Stlengis mesembrinus, описанный недавно из вод »ндонезии. ¬ близком семействе психролютовых (Psychrolutidae) все виды живут за пределами шельфа (общий батиметрический диапазон 150Ч1600 м). Ќекоторые роды (например, Psychrolutes и Cottunculus) имеют бипол€рное распространение.
—емейство липаровых, или морских слизней (Liparididae), очень богатое видами (их более 150), включает как прибрежные формы, обитающие только за пределами теплых вод, так и глубоководные виды, представленные во всех широтных зонах ћирового океана. Ќазвание Ђморские слизниї очень точно характеризует внешний облик этих рыб, голое тело которых имеет студнеобразную консистенцию. »х обычна€ длина составл€ет 10Ч30 см (максимально 70 см). ¬ мезо- и батибентали особенно обыкновенны роды карепроктов (Careproctus) и паралипарисов (Paraliparis), каждый из которых насчитывает примерно по 60 видов. арепрокты обитают на глубине до 3500Ч4000 м, причем шельфовые виды имеют светлую или €рко-розовую окраску, батиальные Ч красную или фиолетовую, а абиссальные Ч почти черную. ¬ соответствии с глубиной обитани€ мен€етс€ также цвет брюшины (от светлой до черной) и диаметр глаз (наибольшего размера они достигают у видов, живущих в сумеречной зоне Ч на глубине 200Ч1000 м). ѕаралипарисы, обитающие в диапазоне глубин от 125 до 3200 м, наиболее многообразны в батиали северной ѕацифики и, как показал ј. ѕ. јндри€шев, в ёжном океане, где находитс€ вторичный центр видообразовани€ группы. —реди липарид имеютс€ даже хадобентальные виды: родственный кapeпроктам псевдолипарис (Pseudoliparis amblystomopsis) обнаружен на глубине около 7600 м в урило- амчатском желобе, а три вида рода Notoliparis в трех разных желобах южного полушари€ ( ермадекском, ёжно-—андвичевом, ћакуори) на глубине более 5000 м.
ћне не приходилось всерьез заниматьс€ липаридами, по все же у мен€ есть одно воспоминание, правда, отнюдь не из при€тных, св€занное с этими рыбами. ¬ 1979 г. Ђ¬ит€зьї обогнул ≈вропу в своем последнем 65-м рейсе, посетив о Ђпрощальным визитомї порты ‘ранции, »спании, ѕортугалии, јнглии и ƒании. ¬ том рейсе принимал участие и ј. ѕ. јндри€шев (мы жили в одной каюте), который очень наде€лс€ использовать посещение јнглии дл€ изучени€ типовых экземпл€ров антарктических паралипари-сов в Ѕританском музее естественной истории. ¬ Ѕискайском заливе нас накрыл жестокий шторм, и јнатолий ѕетрович, поскользнувшись на трапе в момент, когда судно резко повалило на борт, получил очень т€желую травму Ч перелом нескольких ребер, в результате чего оказалс€ в больнице города ƒувра. ќ визите в Ћондон нечего было и думать, и он попросил мен€ проверить некоторые признаки интересовавших его рыб попутно с моим исследованием типового материала по летучкам. ƒл€ работы в музее (€ ездил туда два раза) мне выделили отдельную комнату, куда были доставлены из хранилища банки с отобранными по каталогу экземпл€рами. ѕокончив с летучими рыбами, € открыл сосуд с паралипарисами. “ри сиптипа P. antarcticus - вида, описанного “. –игеном еще в 1912 г., находились в неважном состо€нии: веро€тно, они были исходно фиксированы спиртом, сильно дегидратировались и стали очень твердыми. ѕыта€сь распр€мить одну из рыбок дл€ измерени€, € сделал слишком резкое движение и, о ужас, обломил ее хвост. я оцепенел: изуродовать типовой экземпл€р Ч это непростительное преступление, которому но может быть прощени€. Ќо делать нечего... — повинной головой € направилс€ к ѕитеру ”айтхеду - куратору коллекции рыб и пока€лс€ в своем грехе. Ђј где хвост?ї - спросил ѕитер. Ђ¬от онї. - ЂЌу, ничего. ћы его подклеим так, что будет совсем незаметної. я был прощен, по до сих пор испытываю чувство стыда, вспомина€ это событие.
—амый большой отр€д рыб Ч окунеобразные Ч содержит свыше 6000 видов, но лишь очень немногие из них относ€тс€ к глубоководным обитател€м дна и придонных слоев пелагиали, а на абиссальные глубины вообще проникают считанные формы. —колько-нибудь значительную роль они играют, пожалуй, только в пограничной зоне между шельфом и склоном, где встречаютс€ многие преимущественно шельфовые виды каменных окуней (сем. Serranidae), каталуфовых (Priacanthidae), бланквиловых (Blanchiostegidae), мицуевых (Scombropidae), красноглаз-ковых (Emmelichthyidae), рыб-кабанов (Pentacerotidae) и ƒругих семейств, не опускающиес€, как правило, глубже 400-600 м.
“олько дл€ мезобентопелагиали характерны семейства акропомовых (Acropomatidae) и эпигоновых (Epigonidae), к которым принадлежат рыбы, в преобладающем большинстве живущие в придонных горизонтах на глубине 200Ч600 м. ¬ обоих семействах есть, однако, виды которые насел€ют более глубокие слои (800Ч1100 м), например хауэлла (Howella brodiei), широко распространен па€ во всех океанах, и южный эпигонус (Epigonus robustus), насел€ющий материковые склоны и подводные подн€ти€ в 30Ч40-х широтах южного полушари€. ќдин из наиболее крупных видов Ч эпигонус-телескоп (≈. telescopus), достигающий длины свыше 70 см и массы 4 км также обитает на глубине до 1200 м, где €вл€етс€ объектом промысла. ћальки многих эпигонусов держатс€ в верхнем слое океана.
»нтересно отметить, что в преимущественно эпимезопелагическом семействе морских лещей брамид виды одного рода Ч Eumegistus, €вл€ющегос€, по мнению ƒ. ћида, наименее продвинутым в эволюционном отношении, ведут придонный образ жизни и выход€т в толщу воды только дл€ ночной охоты на миктофид и подобную им по поведению добычу. Ёти лещи достигают довольно крупных размеров (до 70 см) и живут над островными склонами и подводными горами в тропической зоне.
Ѕельдюговые (сем. Zoarcidae) Ч очень многообразна€ холодноводна€ группа северного происхождени€, насчитывающа€ в своем составе около 200 видов, больша€ части которых обитает на глубине 100Ч500 м и даже глубже, хот€ в семействе есть и мелководные, в том числе литоральные, формы. ѕочти все бельдюговые живут непосредственно на дне и т€готеют к м€гким илистым грунтам в которые способны зарыватьс€ сами и закапывать свою икру; питаютс€ они бентосом и донными рыбами. »сключение в этом отношении составл€ют только меланостигмы (род Melanostigma) и ликодапусы (Lycodapus) Ч они могут считатьс€ мезобентопелагическими, так как нередко встречаютс€ в значительном удалении от дна над большими глубинами.( јналогичный образ жизни ведут, по-видимому, представител€ близкого семейства Parabrotulidae Ч мелкие (до 5 см) живоротд€щие рыбки двух родов (Leucobrotula и Parabrotula), ) Ёти рыбы размножаютс€, однако как и все их родичи, откладыва€ крупную донную икру (пр€мых наблюдений нереста, впрочем, не проводилось). ¬ отличие от преобладающего большинства других глубоководных рыб ранние мальки этих и всех остальных зоарцид в верхних сло€х пелагиали не отмечены.
¬ изучение океанических зоарцид, принадлежащих главным образом к подсемейству ликодоподобных (Lycodinae), большой вклад внесен в последние годы ј. ѕ. јндри€шевым, ¬. ¬. ‘едоровым, Ё. јндерсоном. обширному (около 50 видов) роду ликодов (Lycodes) относ€тс€ в основном крупные (длина до 1 м) верхне-мезобентальные формы, и лишь немногие его представители доход€т до батибентали. Ѕлизкие к ликодам, но более миниатюрные лиценхелисы (род Lycenchelys) заметно более глубоководны: эти рыбы полностью отсутствуют па шельфе и вход€т в состав мезо-, бати- и абиссобентальных ихтиоценов. ќба рода богато представлены во всех северных мор€х (они обычны и в наших дальневосточных и пол€рных водах), но встречаютс€ и в южном полушарии, куда их предки проникли с севера по глубинам. ¬ умеренно холодных водах ёжной јмерики они, подн€вшись на шельф, дали начало эндемичным южным родам зоарцид, обитающим и на мелководь€х. числу наиболее глубоководных зоарцид принадлежит и пахикара (Pachycara obesa) (рис. 40), обитающа€ в јтлантическом океане на глубине 2400Ч4800 м. Ѕлизкий род Thermarces был описан совсем недавно –. –озеиблаттом и ƒ. оэном из термальных источников в восточной части “ихого океана на глубине 2600 м. ¬ желудках термарцесов были найдены остатки моллюсков, раков амфипод и вестиментифер рнфтий, существующих за счет сульфобактерий, что позвол€ет считать их посто€нным компонентом этих своеобразных глубоководных сообществ.
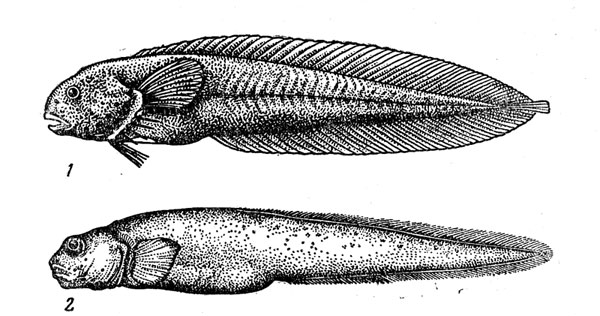
–ис. 40. ѕаралиприс (1) и пахикара (2).
«убатки (два рода семейства Anarhichadidao), распространенные в северных част€х јтлантического и “ихого океанов, живут в основном в шельфовой зоне, но в зимнее врем€ мигрируют на глубины (эта группа всесторонне изучена ¬. ¬. Ѕарсуковым). Ќаиболее глубоководный вид - син€€ зубатка (Anarhichas denticulatus), достигающа€ 1,4 м в длину, отмечена даже на глубине более 900 м. ¬се зубатки откладывают очень крупную донную икру, но их личинки поднимаютс€ в толщу воды, где проход€т весь мальковый период жизни (его продолжительность у разных видов сильно разнитс€).
≈ще одна холодноводна€ группа рыб Ч нототениевидные, эндемична€ дл€ јнтарктики и —убантарктики, где она доминирует. Ёта группа отличаетс€, как показано работами ј. ѕ. јндри€шева, X. ƒевитта, ј. ¬. Ѕалушкина, ј. ¬. Ќеелова, ∆.- . ёро и других ученых, большим систематическим и экологическим разнообразием. ¬ сем. нототениевых (Nototheniidae), усатых дракончиков (Artedidraconidae), антарктических плосконосов (Bathydraconidae) и белокровных рыб (Channichthyidae) есть и глубоководные донно-придонные формы, обитающие на глубине 500Ч1500 м и более. —реди преимущественно прибрежных и шельфовых нототениид наиболее глубоководными €вл€ютс€ антарктическа€ сквама (Lepidonotothen kempi) и черноротый трематом (Trematomus loennbergi), которые держатс€ на склоне континентальной ступени јнтарктиды на глубине 500Ч1000 м, а также патагонский клыкач (Dissоstichus eleginoides) Ч очень крупна€ рыба (длина до 2,1 м, масса до 70 кг), добываемый у островов ёжна€ √еорги€ и ергелен на глубине до 800 м. Ѕольшинство плосконосов живет на глубине, не превышающей 500Ч800 м, но представители рода Bathydraco Ч самого глубоководного среди нототеноидных рыб Ч насел€ют большие глубины (вплоть до 2500 м). ”сатые дракончики рода Pogonophryne также отмечены до глубины 2300 м. √лубинна€ белокровка (Chionobathyscus dewitti), у которой, как и у других ханнихтид, в крови нет ни эритроцитов, ни гемоглобина, также ловилась у берегов ледового континента на глубине до 2000 м. ” всех нототениевидных икра развиваетс€ на дне а личинки ведут пелагический образ жизни.
¬ верхней мезобентали тепловодной зоны (на глубине до 500Ч600 м) вполне обычны также такие рыбы, как хампсодоны (сем. Champsodontidae), звездочеты (Uranoe scopidae: Kathetostoma, Uranoscopus и др.), перкофид (Percophidae, в том числе карликовые виды родов Osopesaron и Pteropsaron, не превышающие 4Ч5 см в длину) драконетки (Draconettidae).

–ис. 41. –уветта (1) а угольна€ рыба-сабл€ (2).
ƒл€ среднеглубинных горизонтов очень характерны саблеподобные рыбы (рис. 41) Ч два близкородственных семейства гемпиловых (Gempylidae) и волосохвостых (Trichiuridae), вход€щие в состав подотр€да скумбриевидных. ¬се они имеют в той или иной степени удлиненное тело, большой рот и очень острые мощные клыкообразиые зубы, легко рассекающие добычу на части. ¬ мезобентопелагических ихтиоценах на глубине 300Ч1000 м (нужно сказать, что эта группа представлена также на шельфе и в океанской пелагиали) особенно важное значение принадлежит прометихтам (род Promethichthys), рекси€м (Rexea), бентодесмам (Benthodesmus) и угольным сабл€м (Aphanopus) Ч хищным рыбам, длина которых нередко достигает 1 м, а также еще более крупной руветте (Ruvettus pretiosus). Ёта рыба может иметь длину 1,8 м при массе около 50 кг. ¬о многих районах мира она служит объектом специального лова на особые снасти, крючки которых опускаютс€ на глубину 300Ч700 м. “акой промысел производитс€, в частности, у берегов убы, ћадейры, оморского архипелага, многих островов ѕолинезии. ћ€со руветты содержит очень много жира, который обладает слабительными свойствами (на некоторых €зыках ее называют Ђкасторковой рыбойї) и при приготовлении рыбы в пищу должен удал€тьс€ во врем€ варки.
—аблеподобным рыбам € посв€тил немало внимани€ и времени. ≈ще в 1955 г., в первом своем плавании в тропиках, мне удалось поймать сачком крупную змеиную макрель, в то врем€ представл€вшуюс€ большой редкостью. ¬ очень попул€рной в те годы книге “ура ’ейердала о путешествии на бальсовом плоту Ђ он “икиї встрече с этим Ђчудищемї посв€щено немало эмоциональных строк. — тех давних пор эта группа посто€нно вызывала мой интерес, и в 1969 г. € предложил ¬ладимиру Ѕеккеру, давнему при€телю (мы вместе учились в институте) и коллеге по лаборатории, совместно разобратьс€ в не очень большой коллекции, которой мы тогда располагали: € вз€лс€ за рыб-сабель, он Ч за гемпилид. ¬ итоге этого внепланового исследовани€ был опубликован предварительный обзор надсемейства, после чего ¬олод€ навсегда вернулс€ к своим любимым миктофидам, а € тоже зан€лс€ другими проблемами, хот€ не тер€л группу из виду и между делом описал в ней еще три вида бентодесм. ¬ середине 70-х € заинтересовал систематикой трихиурид промыслового ихтиолога из ¬Ќ»–ќ —ерге€ ћихайлина, вместе с которым мы дали описани€ двух новых рыб-сабель, но трагическа€ гибель —ережи в конце 1981 г. не позволила ему до конца реализовать свои возможности (он погиб, как герой, спаса€ пассажиров в загоревшемс€ на ходу вагоне поезда алининград - ћосква). ¬ пам€ть о друге (нас объедин€ло не только общее дело, но и одинакова€ страсть к охоте) € назвал его именем один из новых видов угольных сабель, решив поставить на этом точку в изучении этой группы. Ќе получилось... ѕри обработке коллекций с подводного хребта Ќаска вы€снилось, что там обитают два вида рексий - оба неописанные,Ч а исследование сравнительных материалов из других районов “ихого и »ндийского океанов (€ получил их из музеев многих стран) показало, что в роде существуют по меньшей мере шесть видов (считалось, что их только два). –езультаты этой работы еще не опубликованы.
»з океанических строматеевидных рыб к числу придонных принадлежат немногие. Ёто Ч гипероглифы (род Hyperoglyphe) и шедофилы (Schedophilus) из семейства центролофовых Ч крупные рыбы длиной до 1 м, обитающие главным образом в субтропических и умереннотепловодных районах (в том числе над подводными подн€ти€ми) на глубине до 300 м и более. ѕридонно-пелагическими; €вл€ютс€ также ариоммы (около полутора дес€тков видов рода Ariomma), обычные на глубине 100Ч600 м во всех тропических мор€х.
Ќе опускаютс€ глубже мезобентали и преимущественно шельфовые камбалообразные рыбы. “ак, наиболее глубоководпый род ботусовых камбал (сем. Bothidae) Ч хасканопсетты (Chascanopsetta), недавно ревизованный . јмая ока, содержит 5 видов, которые живут на глубине 250-640 м. ќдин из этих видов Ч гавайска€ пеликань€ камбала (Ch. crumenalis) отличаетс€ крайне своеобразным обликом: нижн€€ челюсть у нее очень длинна€ (она на треть: своей длины выступает перед рылом) и поддерживает кожистую горловую сумку, очень напоминающую ту, что имеетс€ у насто€щих пеликанов. ≈ще глубже обитают представители эндемичного дл€ ёжного океана семейства безруких камбал (Achiropsettidae), недавно выделенного —. ј. ≈всеенко и характеризующегос€ отсутствием грудных плавников: максимальна€ глубина вылова неоахиропсетты (Neoachiropsetta milfordl) составл€ет 840 м.
—реди насто€щих, или правосторонних, камбал (сем. Pleuronectidae) есть немало видов, например северные малороты (род Microstomus) или тропические пецилопсетты (Poecilopsetta), приуроченных к пограничной между шельфом и склоном зоне (глубина пор€дка 100Ч 300 м). наиболее глубоководным видам относ€тс€ палтусы, прежде всего черный, или синекорый (Reinhardtius hippoglossoides), и белокорый (Hippoglossus hippoglossus) Ч живущие в северных част€х јтлантического и “ихого океанов хищники, активно преследующие свою добычу в толще воды. ѕолноразмерные особи черного палтуса достигают 1,3 м в длину, но белокорый значительно крупнее (рекордна€ длина 4,7 м при массе 337 кг). —коплени€ всех палтусов обычно приурочены к материковому склону (глубина 150Ч800 м), а наибольша€ глубина их обитани€ превышает километр (черный палтус вылавливалс€ даже па отметке 1700 м). ѕалтусы принадлежат к числу долгоживущих поздносозревающих рыб (белокорый живет до 30 лет, а созревает в массе на 10Ч15-м году). Ќерест происходит в глубоких сло€х воды, личинки сначала тоже держатс€ далеко от поверхности, но, подраста€, перемещаютс€ на мелководь€, где переход€т к донному образу жизни. —реди камбал-плевронектид известны и еще более глубоководные Ч мезобатибентальные виды, например бородавчата€ (Clidoderma asperrimurn) и глубинна€ (≈mbassichlhys bathibius), вполне обычные вплоть до 1800Ч 2000 м от поверхности. ¬ состав батиального ихтиоцена полноправно вход€т и морские €зыки из рода Symphurus (сем. Cynoglossidae), живущие в тропической зоне на глубине 200Ч1000 м. сказанному о камбалообразных нужно еще добавить, что среди глубоководных представителей группы практически нет чистых бентофагов (и уж во вс€ком случае их дол€ значительно ниже, чем среди мелководных).
» наконец, в отр€де скалозубообразных к придонным глубоководным рыбам относитс€ в полном составе семейство холлардиевых (Triacanthodidae), включающее 20 видов, обзор которых сделан недавно ƒ. “айлером. Ёти небольшие (длина не более 25 см) рыбы живут в тропических водах всех океанов на глубине 50Ч600 м и даже глубже. Ѕиологи€ их изучена слабо. —уд€ по некоторым особенност€м, это наименее продвинута€ группа скалозубообразных, но отдельные виды представл€ютс€ очень сильно специализированными. “ак, макрорамфосоды (род Macrorhamphosodes) имеют тонкое рыло, выт€нутое в длинную трубку, на конце которой находитс€ обращенный вбок дисковидный рот. Ёта рыба, как и тидемани€ (Tydemania navigatoris), питаетс€ чешуей других рыб.
—равнение состава бенто-пелагических комплексов разных вертикальных зон (табл. 4) показывает, что число видов в ихтиоценах быстро убывает с глубиной (цифры этой таблицы следует рассматривать как приблизительные). ¬ самой верхней части склона, где представлены многие семейства, типичные дл€ сублиторали, наблюдаетс€ наибольшее разнообразие систематического состава. Ќужно подчеркнуть, что в большинстве случаев речь идет не о случайных проникновени€х отдельных особей тех или иных шельфовых видов в чуждый биотоп, а о посто€нном их обитании в экотонных услови€х у границы смежных вертикальных зон. ¬ справедливости такого утверждени€ убеждает тот факт, что такие виды живут и там, где эпибентальной зоны нет вообще. –ечь идет о подводных подн€ти€х в открытом океане, имеющих минимальные глубины пор€дка 200Ч400 м, т. е. о верхней части талассобатиали в понимании ј. ѕ. јндри€шева. ¬се такие подн€ти€ (их обычно называют Ђбанкамиї) населены тем не менее рыбами, которые более характерны дл€ нижней части шельфа: как правило, их легко сразу опознать по €ркой окраске и совсем не Ђглубоководномуї облику. ¬ качестве типичных эпимезобентальных экотонных рыб можно упом€нуть аулопов (род Aulopus), длиннорылых рыб-солдатов (Ostichthys), антигоний (Antigonia), большеглазых каталуф (Cookeolus), каллантиасов (Callanthias). ¬ерхнемезопелагические виды недавно обнаружены даже в семействах щетинозубовых, или рыб-бабочек (Chaetodontidae), и губановых (Labridae), всегда считавшихс€ приуроченными к прибрежным сообществам, Ч Chaetodon guyotensis и Bodianus thoracotaeniatus описаны с подводных гор хребта юсю-ѕалау (глубина 320Ч340 м).
Ќа сообществах рыб, насел€ющих подводные подн€ти€, нужно остановитьс€ подробнее в св€зи с тем интересом, который они вызывают по многим причинам. ¬ажность их изучени€ первым оценил американский ихтиолог . ’аббз, который еще в 1959 г., основыва€сь на очень скудных фактических данных того времени (фактически на одной случайной поимке триглы на гайоте хребта Ќаска), сформулировал р€д принципиальных вопросов, касающихс€ этой проблемы. ¬от эти вопросы.
ѕосто€нен ли видовой состав населени€ отдельных гор аким образом они были заселены?
Ќе представл€ют ли они опорные пункты дл€ преодолени€ открытоводных барьеров, раздел€ющих прибрежные фауны?
Ќе €вл€ютс€ ли некоторые виды, насел€ющие постепенно погружающиес€ горы, реликтами, адаптированными к увеличенной глубине обитани€?
¬едет ли изол€ци€ гор к видообразованию?
Ќе существует ли на горах и банках и над ними скоплений донных или пелагических рыб и других промысловых объектов?
арл Ћевитт ’аббз (1894-1979) несомненно был одним из самых выдающихс€ ихтиологов нашего столети€. ќн отличалс€ необычайной широтой и глубиной научных интересов и оставил после себ€ весьма достойное наследие - более 700 книг и статей, касающихс€ разнообразнейших аспектов ихтиологии и смежных отраслей знани€. ќн был также простым в общении и доброжелательным человеком с хорошо развитым чувством юмора.
ћне довелось встретитьс€ с . ’аббзом трижды (не могу запросто называть его арлом, как при встречах: по-русски это было бы слишком фамиль€рным и не соответствующим тому уважению, которое € питаю к этому человеку). «накомство состо€лось на √авайских островах в 1964 г. во врем€ очередного “ихоокеанского научного конгресса, на который € попал с Ђ¬ит€земї, совершавшим рейс в центральную ѕацифику. ѕервое общение было очень т€желым как по причине моего несовершенного английского, так и из-за весьма далекого от классических оксфордских норм произношени€ . ’аббза, по € мог с удивлением убедитьс€, что даже в очень узкой специальной области (речь шла о летучих рыбах “ихого океана) он чувствовал себ€ совершенно свободно. Ќа следующий день после моего доклада он шутливо поздравил мен€ с победой в противоборстве Ђс этим варварским английскимї, а через пару мес€цев прислал в ћоскву вырезку из какого-то комикса, в котором говорилось о Ђлетучих коровахї, выдуманных неким впоследствии разоблаченным лжецом, написав на ней: Ђј существуют ли все же летучие рыбы?ї ¬ 1968 г. Ђјкадемик урчатовї зашел в ¬иллемстад (о-в юрасао), когда там происходила международна€ конференци€ по арибскому морю, в которой, оказалось, принимал участие и . ’аббз. ќн был очень подвижек и каждый год предпринимал дальние во€жи вместе со своей верной женой и помощницей Ћорой ’аббз. Ќа конференции он заметил мен€ первым (€, надо сказать, совсем не ожидал этой встречи), сразу узнал и с такой непринужденностью Ђпроехалс€ї насчет моей изменившейс€ внешности (€ стал носить бороду), что два молодых ихтиолога с Ѕарбадоса, с которыми € беседовал, с полной уверенностью сочли его моим бывшим профессором. ѕоследний раз € видел ’аббза в 1979 г. в алифорнии. ќн был болен и знал, что его дни сочтены, сильно ослабел и постарел, но продолжал трудитьс€, интересовалс€ работой коллег, говорил о желании прин€ть участие в следующем “ихоокеанском конгрессе в ’абаровске... ѕрах арла ’аббза по его завещанию был разве€н над “ихим океаном. Ѕольшой фотопортрет с надписью Ђ...советскому коллеге от старого мор€каї, запечатлевший его сто€щим в полный рост на корме штормующего в океане судна, напоминает мне о наших встречах.
“аблица 4
| ќтр€д, семейство | ¬сего видов | ¬идов в ихтиоценах | ¬стречаемость в других ихтиоценах | |||||||
| Ёћ | ћ | ћЅ | Ѕ | Ѕј | ј | ј’ | ’ | |||
| ћиксинообразные (Myxiniformes) | ||||||||||
| ћиксиновые (Myxinidae) | 32 | 20 | 6 | 1 | - | - | - | - | - | Ёпибенталь |
| ћногожаберникообразные (Hexanchiformes) | ||||||||||
| ѕлащеносные акулы (Chlamydoselachidae) | 1 | - | 1 | - | - | - | - | - | - | Ќет |
| атранообразные (Squalijormes) | ||||||||||
| атрановые акулы (Squalidae) | 68 | 12 | 38 | 8 | - | 1 | - | - | - | Ёпимезопелагиаль |
| ÷ентриновые акулы (Oxynotidae) | 4 | 2 | 2 | - | - | - | - | - | - | Ёпимезопелагиаль |
| Ћамнообразные (Lamniformes) | ||||||||||
| ѕесчаные акулы (Odontaspidae) | 4 | 1 | 1 | - | - | - | - | - | - | Ёпибенталь |
| јкулы-домовые (Mitsukuriuidae) | 1 | - | 1 | - | - | - | - | - | - | Ќет |
| архаринообразные (—archarhiniformes) | ||||||||||
| ошачьи акулы (Scyliorhinidae) | 86 | 13 | 45 | 4 | - | - | - | - | - | Ёпибенталь |
| ѕросциллиевые (Proscyllidae) | 6 | 3 | 1 | - | - | - | - | - | - | " |
| Ћожнокуньи акулы (Pseudotriakidae) | 1 | - | 1 | - | - | - | - | - | - | Ќет |
| уньи акулы (Triakidae) | 34 | 5 | 1 | - | - | - | - | - | - | Ёпибенталь |
| √нюсообразные (Torpediniformes) | ||||||||||
| √нюсовые скаты (Torpedinidae) | 15 | 5 | 1 | - | - | - | - | - | - | " |
| Ќарковые скаты (Narkidae) | 25 | 6 | 2 | - | - | - | - | - | - | " |
| —катообразные (Rajiformes) | ||||||||||
| –омбовые скаты (Rajidao) | 200 | 70 | 65 | 15 | - | - | - | - | - | Ёпибенталь |
| Ћожноромбовые скаты (Pseudorajidae) | 4 | - | 4 | - | - | - | - | - | - | Ќет |
| Ќитерылые скаты (Anacanthohatidae) | 15 | 2 | 13 | - | - | - | - | - | - | " |
| Xвостоколообразные (Myliobatiformes) | ||||||||||
| Ўестижаберные скаты (Hexatrygonidae) | 3 | - | 3 | - | - | - | - | - | - | Ќет |
| ’имераобразные (Chimaeriformes) | ||||||||||
| ’имеровые (Chimaeridae) | 20 | 2 | 18 | - | - | - | - | - | - | " |
| Ќосатые химеры (Rhinochimaeridae) | 6 | - | 4 | 2 | - | - | - | - | - | " |
| —пиношипообразные (Notacanthiiormes) | ||||||||||
| √алозавровые (Halosauridae) | 15 | - | 4 | 9 | 1 | 1 | - | - | - | " |
| —пиношиповые (Notacanthidae) | 9 | 2 | 3 | 4 | - | - | - | - | - | " |
| Ћипогеновые (Lipogenydae) | 1 | - | 1 | - | - | - | - | - | - | " |
| ”греобразные ( Anguilliformes) | —литножаберные угри (Synaphobranchidae) | 28 | 2 | 7 | 4 | 1 | 6 | - | - | - | Ёпибенталь |
| Ќеттастомовые (Nettastomatidae) | 35 | 4 | 12 | 4 | - | - | - | - | - | " |
| онгеровые (Congridae) | 110 | 20 | 20 | - | - | - | - | - | - | " |
| ћоногнатовые (Monognathidae) | 14 | - | 3 | 2 | 4 | 2 | 3 | - | - | Ќет |
| Ћососеобразные (Salmoniformes) | ||||||||||
| —еребр€нковые (Argentinidae) | 35 | 15 | 7 | - | - | - | - | - | - | ћезопелагиаль |
| √ладкоголововые (Alepocephalidae) | 98 | 3 | 40 | 30 | 20 | 5 | - | - | - | Ќет |
| Ћептохиловые (Leptochilichthyidae) | 2 | - | 1 | - | - | 1 | - | - | - | " |
| ѕлатитроктовые (Platytroctidae) | 40 | 1 | 27 | 10 | 2 | - | - | - | - | " |
| —томиеобразные (Stomiiformes) | ||||||||||
| √оностомовые (Gonostomatidae) | 26 | - | 3 | - | - | - | - | - | - | ћезобатипелагиаль |
| “опориковые (Sternoptychidae) | 51 | - | 37 | - | - | - | - | - | - | ћезобатипелагиаль |
| ‘отихтовые (Photichthyidae) | 21 | - | 4 | - | - | - | - | - | - | " |
| јстронестовые (Astronesthidae) | ||||||||||
| ћеланостомиевые (Melanostomiatidae) | 27 | - | 5 | - | - | - | - | - | - | ћезопелагиаль |
| Ayлапообразные (Aulopiformes) | ||||||||||
| јулоповые (Aulopidae) | 10 | 5 | 1 | - | - | - | - | - | - | Ёпибенталь |
| «еленоглазковые (Chlorophthalmidae) | 45 | 20 | 7 | - | - | - | - | - | - | Ќет |
| Ќотосудиевые (Notosudidae) | 19 | - | 17 | - | - | - | - | - | - | ћезопелагиаль |
| ящероголововые (Synodontidae) | 40 | 6 | 1 | - | - | - | - | - | - | Ёпибенталь |
| ћиктофообразные (Myctophiformes) | ||||||||||
| Ќеоскопеловые (Neoscopelidae) | 5 | - | 4 | - | - | - | - | - | - | ћезопелагиаль |
| ћиктофовые (Myctophidae) | 220 | 8 | 12 | - | - | - | - | - | - | ћезобатипелагиаль |
| “рескообразные (Gadiformes) | ||||||||||
| ѕаркетниковые (Muraenolepididae) | 4 | 1 | 1 | - | - | - | - | - | - | Ёпибенталь |
| ћоровые (Moridae) | 90 | 20 | 40 | 15 | - | - | - | - | - | " |
| ћеланоновые (Melanonidae) | 2 | - | 1 | - | - | - | - | - | - | ћезопелагиаль |
| Ѕрегмацеровые (Bregmacerotidae) | 7 | - | 5 | - | - | - | - | - | - | " |
| “ресковые (Gadidae) | 50 | 12 | 3 | - | - | - | - | - | - | Ёпибенталь |
| ћерлузовые (Merlucciidae) | 25 | 24 | 1 | - | - | - | - | - | - | Ќет |
| ƒолгохвостовые (Macrouridae) | 350 | 10 | 245 | 45 | 15 | 30 | 5 | - | - | " |
| ќшибнеобразные (Ophldiiformes) | ||||||||||
| ќшибневые (Ophidiidae) | 170 | 15 | 35 | 30 | 20 | 30 | 12 | 3 | - | Ёпибенталь, мезопелагиаль |
| Ѕититовые (Bythitidae) | 85 | 13 | 13 | 3 | 2 | 2 | - | - | - | Ёпибенталь |
| јфионовые (Aphyonidae) | 18 | - | 5 | - | 2 | 3 | 8 | - | - | Ќет |
| ”дильщикообразные (Lophiiformes) | ||||||||||
| ”дильщиковые (Lopliiidae) | ||||||||||
| ”дильщиковые (Lopliiidae) | 25 | 12 | 8 | 1 | - | - | - | - | - | Ёпибенталь |
| ’аунаксовые (Chaunacidae) | 10 | 2 | 8 | - | - | - | - | - | - | Ќет |
| Ќетопыревые (Ogcocephalidae) | 60 | 12 | 26 | 7 | - | - | 1 | - | - | Ёпибенталь |
| “ауматихтовые (Thaumatichthyidae) | 5 | - | - | - | - | - | 1 | - | - | ћезобатипелагиаль |
| ƒицерациевые (Diceratiidae) | 5 | - | - | 5 | - | - | - | - | - | Ќет |
| итовидкообразные (Cetomimiformes) | ||||||||||
| јтелеоповые (Ateleopidae) | 12 | 3 | 9 | - | - | - | - | - | - | " |
| Ѕериксообразные (Berycidae) | ||||||||||
| “рахихтовые (Trachichthyidae) | 38 | 11 | 21 | - | - | - | - | - | - | Ёпибенталь |
| ƒиретмовые (Diretmidae) | 4 | - | 1 | 2 | - | - | - | - | - | ћезопелагиаль |
| Ѕериксовые (Berycidae) | 8 | 7 | 1 | - | - | - | - | - | - | Ќет |
| —тефанобериксовые (Stephanoberycidae) | 3 | - | - | 1 | - | 2 | - | - | - | " |
| ћеламфаевые (Melamphaidae) | 33 | - | 2 | - | - | - | - | - | - | ћезабатипелагиаль |
| √испидобериксовые (Hispidoberycidae) | 1 | - | 1 | - | - | - | - | - | - | Ќет |
| √ибберихтовые (Gibberichthyidae) | 2 | - | - | 2 | - | - | - | - | - | " |
| —олнечникообразные (Zeiformes) | ||||||||||
| ѕаразеновые (Parazentidae) | 1 | - | 1 | - | - | - | - | - | - | " |
| «енионовые (Zeniontidae) | 4 | - | 4 | - | - | - | - | - | - | Ќет |
| ÷иттомимовые (Cyttomimidae) | 2 | - | 2 | - | - | - | - | - | - | " |
| Ћунниковые (Oreosomatidae) | 10 | - | 8 | 2 | - | - | - | - | - | " |
| „ешуеигловые (Grammicolepididae) | 4 | - | 4 | - | - | - | - | - | - | " |
| апровые (Caproidae) | 11 | 9 | 2 | - | - | - | - | - | - | " |
| —корпенообразные (Scorpaentformes) | ||||||||||
| —корпеновые (Scorpaenidae) | 310 | 80 | 15 | 1 | - | - | - | - | - | Ёпибенталь |
| ѕеристедиевые (Periostediontidae) | 25 | 10 | 10 | - | - | - | - | - | - | " |
| ѕлоскоголововые (Platycephalidae) | 60 | - | 1 | - | - | - | - | - | - | " |
| јноплопомовые (Anoplopomatidae) | 2 | 2 | - | 1 | - | - | - | - | - | Ќет |
| –огатковые (Cottidae) | 300 | 45 | 10 | - | - | - | - | - | - | Ёпибенталь |
| ѕсихролютовые (Psychrolutidae) | 30 | 10 | 10 | 5 | - | - | - | - | - | " |
| Ћипаровые (Liparididae) | 150 | 25 | 60 | 25 | 12 | 5 | - | - | - | Ёпибенталь |
| ќкунеобразные (Perciformes) | ||||||||||
| Ёпигоновые (Epigonidae) | 26 | 3 | 22 | 1 | - | - | - | - | - | Ќет |
| ћорские лещи (Bramidae) | 17 | - | 2 | - | - | - | - | - | - | Ёпипелагиаль |
| Ћжесельдевые (Bathyclupeidae) | 4 | - | 3 | 1 | - | - | - | - | - | Ќет |
| ўетинозубовые (Chaetodontidae) | 114 | - | 1 | - | - | - | - | - | - | Ёпибенталь |
| абан-рыбы (Pentacerotidae) | 12 | 5 | 3 | - | - | - | - | - | - | " |
| ÷еполовые (Cepolidae) | 20 | 9 | 1 | - | - | - | - | - | - | " |
| √убановые (Labridae) | 500 | 5 | 1 | - | - | - | - | - | - | " |
| Ѕельдюговые (Zoarcidae) | 160 | 50 | 45 | 20 | 5 | 3 | 7 | - | - | " |
| Ќототениевые (Nototheniidae) | 50 | 30 | 2 | - | - | - | - | - | - | " |
| ”сатые дракончики | 15 | 8 | - | 1 | - | - | - | - | - | " |
| Ѕатидраковые (Bathydraconidae) | 16 | 3 | 2 | 3 | 1 | - | - | - | - | Ќет |
| Ѕелокровные рыбы (Channichthyidae) | 16 | 6 | 4 | 4 | 1 | - | - | - | - | Ёпибенталь |
| ѕеркофисовые (Percophidae) | 35 | 15 | 15 | - | - | - | - | - | - | " |
| ƒраконетковые (Draconettidae) | 10 | 3 | 7 | - | - | - | - | - | - | Ќет |
| √емпиловые (Gempylidae) | 22 | 3 | 13 | - | - | - | - | - | - | Ёпимезопелагиаль |
| —абли-рыбы (Trichiuridae) | 27 | 3 | 21 | - | - | - | - | - | - | Ёпибенталь |
| ÷ентролофовые (Centrolophidae) | 22 | 9 | 4 | - | - | - | - | - | - | Ёпипелагиаль |
| амбалообразные (Pleuronectiformes) | ||||||||||
| Ѕотусовые (Bothidae) | 210 | 20 | 15 | - | - | - | - | - | - | Ёпибенталь |
| амбаловые (Pleuronectidae) | 100 | 49 | 12 | - | - | - | - | - | - | Ёпибенталь |
| ÷иноглоссовые (Cynoglossidae) | 100 | 30 | 20 | - | - | - | - | - | - | " |
| —олевые (Soleidae) | 120 | 15 | 1 | - | - | - | - | - | - | " |
| —калозубообразные (Tetraodontiformes) | ||||||||||
| ’оллардиевые (Triacanthodidae) | 20 | 10 | 10 | - | - | - | - | - | - | Ќет |
(ѕримечание. ¬ таблицу не включены отр€ды и семейства, представленные только в переходной от шельфа к склону зоне (Ёћ) Ч миногообразные (Petromyzontiforrnes): миноговые (Petrornyzontidae) Ч 1 вид; многожаберникообразные: многожаберные акулы (Hexanchidae) Ч 4; катранообразные: звездчатошипые акулы (Echinorhinidae) Ч 2; плоскотелообразные (Spuatini-formes): плоскотелые акулы (Squatinidae) Ч 2; кархаринообразные: серые акулы (Carcharhinidae) Ч 1; скатообразные: новозеландские скаты (Arhynchobatidae) Ч 1: целакантообразные (Coelacanthiformes): латимерисвые (Latimeriidae) Ч 1; тарпонообразные (Elopjformes): гисовые (Pterothrissidae) Ч 2j угреобразные: ложнококгеровые (Xenocongridae) Ч 1, острохвостые угри (Ophichthidae) Ч 5. макроцефенхеловые (Macrocephenchelyidae) Ч 1: ошибнеобразные: карапусовые (Carapidae) Ч 5: удильщикообразные: клоуновые (Antennariidae) Ч 3; бериксообразаые: шишечниковые (Monocentridae) Ч 4, голоцентровые (Holocentridae) Ч 12, полимиксиевые (Polymixiidae) Ч 6; солнечникообразные: солнечниковые (Zeidae) Ч 10; колюшкообразные (Gasterosteiformes): бекасовые (Macrorhamphosidae) Ч 8; скорпенообразные: конгиоподовые (Congiopodidae) Ч 5, тригловые (Triglidae) Ч 15, гоплихто-вые (Hoplichthyidae) Ч 10, терпуговые (Hexagrammidae) Ч 2, эреуниевые (Ereuniidae) Ч 2, лисичковые (Agontdae) Ч 15; окунеобразные: каменные окуни (Serranidae) Ч 40, полиприоновые (Polyprionidae) Ч 3, каллантиевые (Callanthiidae) Ч 10, приакантовые (Priacanthidae) Ч 5, акропомовые (Acropomatidae) Ч 15, мицуевые (Scombropidae) Ч 4, малакантовые (Malacanthidae) Ч 20, ставридовые (Carangidae) Ч 6, красноглазковые (Emmelichthyidae) Ч 11, луциановые (Lutianidae) Ч 10, спаровые (Sparidae) Ч 15, центракантовые (Gentracanthldae) Ч 2, джакасовые (Cheilodactylidae) Ч 5, трубачевые (Latriobidae) Ч 1, стихеевые (Stichaeidae) Ч 10, зубатковые (Anarhichadidae) Ч 4, хампсодоновые (Champsodontidae) Ч 6, волосозубовые (Trichodontidae) Ч 2, звездочетовые (Uranoscopidae) Ч 5, мугилоидовые (Mugiloididae) Ч 5, пескарковые (Callyonimidae) Ч 15, бычковые (Gobiidae) Ч 10, ариоммовые (Ariommidae) Ч 10; камбалообразные: безрукие камбалы (Achiropsettidae) Ч 12; скалозубообразные: триодоновые (Triodontidae)Ч 1. Ёти 345 видов включены в итоговую строку колонки Ёћ.)
»зучение рыб подводных подн€тий только начинаетс€, но уже сейчас можно отметить, что в разных районах ћирового океана в заселении этого биотопа прин€ли участие в основном представители одних и тех же, притом сравнительно немногих, семейств. ак показывает сравнение ихтиофаунистических списков подводных гор ћетеор и ∆озефина в северо-восточной јтлантике, банки ¬альдиви€ итового хребта в юго-восточной јтлантике, тихоокеанских подводных хребтов юсю-ѕалау, √авайского, —еверо-«ападного, Ќаска и —ала-и-√омес, сходство в составе населени€ этих далеко отсто€щих друг от друга участков довольно велико, особенно на уровне семейств и родов.
¬идовой состав ихтиоценов на подводных подн€ти€х, определ€етс€ в первую очередь глубиной над их вершинами, а разнообразие этого состава Ч действием Ђостровного эффектаї, пр€мо завис€щего от площади подн€ти€ “алассоэпибентальные ихтиоцены на вершинах самых мелководных банок (например, гор елсо, ейпл и ”анганелла в “асмановом море, имеющих глубину 40Ч100 м) обычно представл€ют собой в той или иной степени обедненный вариант сообществ ближайших шельфовых районов. Ќе исключена возможность, что на крохотных вершинных площадках посто€нные самовоспроизвод€щиес€ попул€ции эпибентальных рыб вообще отсутствуют и их население пополн€етс€ только за счет оседани€ пелагических личинок, принесенных извне, т. е. с ближайшего шельфа.
—уществование независимых горных попул€ций рыб эпимезо- и мезобентопелагического комплекса, напротив, не вызывает никаких сомнений. ќб этом свидетельствуют не только многолетние наблюдени€ над локальными попул€ци€ми таких, к примеру, видов, как рыба-кабан Pseudopentaceros wheeleri на √авайском подводном хребте, капродон Caprodon longimanus на уже упоминавшейс€ банке ”анганелла, безпузырный окунь Helicolenus lengerichi и зеркальный солнечник Zenopsis sp. (неописанный вид, близкий к Z. nebulosus) на подводном хребте Ќаска, но и существование эндемичных видов, ареалы которых ограничены одним подн€тием или группой близкорасположенных подн€тий. Ёндемичные талассобатиальные виды известны сейчас из многих районов ћирового океана, в том числе с гор јзорского архипелага и с итового хребта в јтлантическом океане, ћадагаскарского и ¬осточно-»ндийского хребтов в »ндийском океане, хребтов юсю-ѕалау, —еверо-«ападного, Ќаска и —ала-и-√омес в “ихом океане. ѕоследний район Ч хребты тепловодной юго-восточной ѕацифики Ч представл€етс€, кстати сказать, прекрасным местом дл€ разнопланового модельного изучени€ талассных ихтиоценов, в том числе и дл€ ответа на вопросы, поставленные . ’аббзом. ”никальность этих хребтов состоит в том, что, несмотр€ на близость к материку ёжной јмерики, они заселены рыбами индовестпацифического происхождени€, и поэтому ближайшие шельфы и склоны €вно не поставл€ют никакого материала дл€ пополнени€ их населени€. —ами эти горы представл€ют собой погруженные коралловые острова (гайоты), различающиес€ по глубине и возрасту (геологическому времени существовани€).
¬есной 1980 г. в нашу лабораторию поступили материалы, несколькими мес€цами раньше собранные моими бывшими аспирантами √. ј. √олованем (теперь он возглавл€ет научные работы в —евастопольском бюро по подводным исследовани€м) и Ќ. ѕ. ѕахоруковым. ќни принимали участие в рейсе Ђ»хтиандраї на подводные хребты Ќаска и —ала-и-√омес, которые до того были совсем неизученными в ихтиологическом отношении (правда, еще раньше там работали сотрудники “»Ќ–ќ, но, к большому сожалению, они, как и во многих других рыбохоз€йственных экспедици€х, не собирали представительных коллекций рыб). ѕредварительна€ обработка полученных нами проб (ее проводили ё. —азонов, ё. ўербачев и €) дала на редкость интересные и, € бы сказал, неожиданные результаты. ¬ы€снилось, что среди донных и придонных рыб этого района восточной ѕацифики преобладают виды, которые были известны только из западной и центральное частей океана Ч от японии, јвстралии, √авайских островов, Ќовой аледонии, а многие другие относ€тс€ к числу неописанных; ¬последствии состо€лось еще несколько экспедиций в этот ра€ он - € сам побывал там дважды на Ђѕрофессоре ћес€цевеї и Ђѕрофессоре Ўтокмаыеї и в результате почти полностью переключилс€ на изучение Ђгорныхї рыб. —ейчас список видов, обнаружейных на обоих хребтах, увеличилс€ до 160, и почти треть из них Ч это исключительно высокий процент Ч оказалась эндемичной (описани€ 30 новых видов уже опубликованы, остальные готов€тс€ к печати). ¬ моих ближайших планах одно из первых мест занимает сейчас подготовка монографии Ђ–ыбы подводных хребтов Ќаска и —ала-и-√омесї, над которой мы работаем вмести с сотрудником ¬Ќ»–ќ ј. Ќ. отл€ром (у него € тоже был когда-то руководителем по кандидатской диссертации), наде€сь завершить ее через год Ч другой.
» еще: не могу не упом€нуть о трудност€х тралений н гоpax. ѕредставление об этих трудност€х легко можно получить вообразив, что нужно провести трал с дирижабл€, лет€щего выше сплошного облачного покрова, по склонам, скажем, авказского хребта. Ёта мысль посто€нно приходит мне в голову, когда € смотрю на горы в иллюминатор самолета, и такое действие представл€етс€ просто неверо€тным, но тем не менее... ѕри первых в моей практике работах на подводных подн€ти€х (это был горный массив Ёкватор в западной части »ндийского океана) мы раз за разом опускали с Ђ–ифтаї оттертрал на склоны (естественно, не вслепую, а ориентиру€сь по эхолоту), порвали все, что рветс€ но так и не смогли получить полноценной пробы. ќсобенно запомнилс€ один задев, когда тралова€ доска, попавша€ в расщелину, превратилась у нас в мертвый €корь и вышедшее на верти кальный трос (мор€ки говор€т Ђна панерї) и содрогающеес€ oт напр€жени€ судно не могло на любом курсе сдвинутьс€ с местаЋ пока 25-миллиметровый стальной ваер не лопнул с таким громоподобным звуком, что создалось полное впечатление взрыва. ’орошо работать только на ровных вершинах гайотов, да и там трал могут подстерегать разнообразные и неожиданные сюрпризы —реди рыб, составл€ющих посто€нное население вершин подводных гор, есть немало чисто донных видов (в том числе зеленоглазки, хаунаксы, скорпениды, морские петухи, камбалы), но наиболее массовый и характерпый компонент сообщества составл€ют бентопелагические объекты Ч колючие акулы, мавролик, специфичные дл€ гор рыбы-топорики и свет€щиес€ анчоусы, рыбы-бекасы мориды, долгохвосты, солнечники, бериксы, большеголовы, некоторые ставридовые, каменные окуни, эпигонусы, красноглазки, рыбы-сабли, масл€ные рыбы и др.
Ѕиологи€ большинства Ђгорныхї бентопелагических рыб пока изучена слабо. »звестно, правда, что многие из них совершают суточные вертикальные миграции: днем они держатс€ вблизи дна, а в темное врем€ поднимаютс€ в верхнюю толщу вод, удал€€сь на 100Ч200 м и более от твердой поверхности. ак уже указывалось, такие миграции предпринимают даже мелкие стайные виды (Maurolicus muelleri, Diaphus suborbitalis). ¬ св€зи с этим возникает естественный вопрос, как удаетс€ этим рыбам, несмотр€ на наличие течений, иногда достаточно сильных, всю ночь оставатьс€ над подн€тием, с которым они тер€ют визуальный контакт, и точно опускатьс€ на него на рассвете. ¬се объ€снени€ этого феномена (ориентаци€ по светилам, пол€ризованному свету, градиентам скорости и т. п.) чисто умозрительны и выгл€д€т неубедительно.
¬озможность массовых миграций талассобатиальных рыб от одного подводного подн€ти€ к другому кажетс€ мне весьма проблематичной, если не говорить о горах, расположенных на одном цоколе: в этом случае эврибатные виды (низкотелый берикс, например, обитает в диапазоне от 50 до 1000 м), естественно, могут перемещатьс€ в пределах всего района, подход€щего по глубине. Ёта точка зрени€ не €вл€етс€, однако, общеприн€той. ѕредполагают, в частности, что рыба-кабан √авайского подводного хребта проводит от 4Ч5 до 7 лет в пелагиали северной ѕацифики и только после этого приходит на горы, где размножаетс€. ¬есьма логична€ схема функциональной структуры попу-л€ционных ареалов берикса в јтлантическом океане, в основу которой положены данные о размерно-возрастном составе и зрелости рыб в разных районах, была недавно разработана ‘. ≈. јлексеевым с соавторами. ¬от как описываетс€, например, цикл существовани€ североатлантической попул€ции: нерестова€ часть ареала приурочена к горам ”глового подн€ти€ в западной части круговорота; оттуда пелагические икра, личинки и мальки дрейфуют в —еверо-јтлантическом течении на восток и юго-восток; молодь оседает на подводных подн€ти€х в восточной части круговорота, а в отдельные годы и на склоне северозападной јфрики; по мере полового созревани€ берикс активно мигрирует на запад по южной периферии круго-ворота, и половозрелый контингент посто€нно обитает в –епродуктивной области (т. е. на ”гловом подн€тии). я думаю, что региональные различи€ в размерах берикса, вз€того из промысловых уловов, могут объ€сн€тьс€ разное глубиной лова (половозрелые особи обитают глубже, чем созревающие, а траловые участки на ”гловом подн€тии наход€тс€ на большей глубине, чем на јзорских банках) Ѕольшое сомнение вызывает и сама способность мигрантов к отысканию нерестовых участков Ч дл€ этого нужно предположить почти сверхъестественные навигационные cпoсобности рыб. » наконец, этому представлению, как и всем основанным на косвенных доказательствах соображени€м в пользу существовани€ таких миграций, противоречит полное отсутствие этих рыб над большими глубинами в открытом океане. ƒо сих пор не отмечено ни одной; поимки взрослых бериксов, красноглазок и подобных им объектов в далеком удалении от материковых склонов подводных возвышенностей.
’уже всего изучены жизненные циклы бентопелагических рыб и особенно их размножение и ранние стадии онтогенеза, во многом загадочные в св€зи с исключительно редкой встречаемостью личинок и мальков в уловах, полученных любыми оруди€ми лова. ќтносительно воспроизводства рыб, обитающих на подводных подн€ти€х, высказывались два предположени€. —огласно одной точке зрени€, развитие их планктонных стадий происходит системах местных циркул€ции вблизи гор. јльтернативна€ гипотеза о заселении подн€тий личинками и мальками, приносимыми течени€ми Ђизвнеї, также имеет своих сторонников.
ƒействительно, слаба€ изученность процессов воспроизводства талассобентальных рыб сильно преп€тствует однозначному решению вопроса. ¬ то же врем€ доказанпость существовани€ независимых попул€ций на отдельных банках, например на горе ћетеор в јтлантическом; океане или на некоторых группах близкорасположенных подн€тий хребтов Ќаска и —ала-и-√омес, дает полное ocнование отказатьс€ от представлени€ об основополагающей роли разноса икры, личинок и мальков течени€ми и наличии значительного обмена генофондами. –азвити€ планктонных стадий может, по-видимому, происходите в непосредственной близости от подводных возвышенностей в локальных круговоротах любого пространственного масштаба, даже не имеющих стационарного характера, но обладающих такой временной устойчивостью, котора€ была бы соизмерима с продолжительностью пассивного пребывани€ икры и личинок в пелагиали (около 10Ч15 суток). ≈ще одна гипотетическа€ возможность, требующа€ экспериментальной проверки, состоит в откладывании икринок с отрицательной плавучестью в щели и углублени€ дна. Ѕолее того, как показало исследование разработанной ¬. Ѕ. ÷ейтлиным и ё. ј. –уд€ковым имитационной модели независимой попул€ции рыб, насел€ющих подводную гору, така€ попул€ци€ вполне способна сохран€тьс€ и воспроизводитьс€ даже и при разносе рыбной молоди на планктонных стади€х развити€ течени€ми, если благодар€ турбулентной диффузии она будет иметь шанс случайно вернутьс€ к месту рождени€. »з этой модели следует, правда, что вниз по течению от горы должен выт€гиватьс€ довольно значительный Ђшлейфї икры и личинок, чего в природе никогда не наблюдали.
»так, что же про€снилось в наших знани€х о рыбах подводных подн€тий за три дес€тка лет, прошедших с того времени, когда арл ’аббз задал свои вопросы? —тало €сно, что видовой состав ихтиоценов отдельных подн€тий характеризуетс€ значительным посто€нством во времени, ибо кажда€ гора представл€ет собой особый биотоп с насел€ющим его биоценозом. «аселение гор происходило точно тем же путем, что и заселение островов и их цоколей Ч с шельфов и склонов материков, и поэтому подводные подн€ти€ должны рассматриватьс€ в том же р€ду природных объектов, что и океанические острова. олонизаци€ островных биотопов в открытом океане (включа€ горы) шла в основном за счет более разнообразных прибрежных фаун, насел€ющих западные части океанов, и чем дальше расположено подн€тие от главного источника своего формировани€, тем выше эндемизм его населени€ (максимальна€ дол€ эндемиков Ч около 40% в диапазоне глубин 200Ч500 м вы€влена на хребтах Ќаска и —ала-и-√омес). ѕодн€ти€, конечно же, €вл€ютс€ Ђопорными пунктамиї в расселении тех рыб, которые способны к достаточно долгому выживанию в пелагиали во взрослом состо€нии или на стадии молоди (хороший пример дают поимки в пелагиали восточной тропической ѕацифики крупных мальков большеглазой каталуфы Cookeolus boops, €вно происход€щих от попул€ций подводных гор). “акже очевидно, что среди среднеглубинных видов есть потомки мелководных форм (уже упоминавшийс€ щетинозуб Chaetodon guyotensis с хребта юсю-ѕалау Ч типичный дериват фауны коралловых рифов), возникшие в результате заглублени€ вершин подн€тий. Ќе вызывает никаких сомнений и вполне очевидный сейчас факт видовой дивергенции рыб в услови€х изол€ции на горах, имеющих достаточно длительную историю существовани€. ¬опрос о скоплени€х рыб у подводных подн€тий тоже не осталс€ без ответа (этот аспект обсуждаетс€ в гл. VI). “аким образом, все Ђзагадкиї . ’аббза вроде бы разгаданы, но полной €сности в талассобатиальной проблеме все еще нет: на передний план выдвинулись теперь экологические aспекты существовани€ горных попул€ций. Ѕудущие ихтиологические изыскани€ на океанских Ђбанкахї должны быть направлены прежде всего на изучение размножени€, жизненных циклов, миграций и особенностей поведени€ рыб (здесь никак нельз€ обойтись без использовани€ обитаемых подводных аппаратов), а также на вы€вление попул€ционной структуры отдельных видов.
Ѕиотопическое своеобразие подводных подн€тий, о котором шла речь выше, не может затушевать, однако, очевидного сходства их населени€ с населением материковых склонов в тех же диапазонах глубин. ѕовсюду сама€ верхн€€ часть среднеглубинной зоны имеет переходный (иначе говор€, экотонный) характер, по батиметрические границы экотона и степень его своеобрази€, которую можно выразить процентом видового эндемизма в пределах эпимезобентопелагического комплекса рыб, насчитывающего более 1100 видов (см. табл. 4), сильно варьируют в разных участках ћирового океана.
»хтиоцены талассной мезобентали отличаютс€, насколько можно судить по имеющимс€ данным, главным образом заметным видовым обеднением по сравнению с прилегающими участками континентальных склонов. Ќа итовом хребте, например, на глубине 200Ч1500 м отмечено всего 60Ч70 видов рыб, что в 3Ч4 раза меньше, чем на соответствующих глубинах склона юго-западной јфрики. ѕо ё. Ќ. ўербачеву, из 36 видов батиальных офидиид »ндийского океана 20 найдены пока только на материковом склоне и лишь 3 только в талассобатиали (13 видов €вл€ютс€ общими). Ёто обеднение св€зано не только с отсутствием видов, проникающих в мезобеиталь с шельфа, но, по-видимому, отражает и меньшую Ђемкостьї талассобатиали Ч меньшее разнообразие экологических ниш.
ћезобенталь и прилегающа€ к ней водна€ толща заселены хорошо обособленным сообществом рыб, в состав которого входит почти 1200 специфических видов. ѕравда, число эндемичных дл€ этого биотопа семейств невелико, по к нему приурочено наибольшее разнообразие многих групп. ќсобенно характерны дл€ этой зоны колючие акулы, в частности из рода Etmcpterus, кошачьи акулы Apristurus, ромбовые скаты, слитиожаберыые угри, гладкоголовы, моровые, долгохвосты (наиболее богата€ видами группа донно-придоиных глубоководных рыб), ошибневые, бериксообразпые, солнечиикообразные и др.
Ѕольша€ часть рыб, насел€ющих верхнюю часть склона на глубине менее 1500 м, принадлежит к придонным и донным видам, однако среди них есть, как уже говорилось, и чисто пелагические формы, не имеющие непосредственной св€зи с дном и в какой-то степени аналогичные неритическим рыбам эшгаелагиали. ѕомимо насто€щих пектонных рыб Ч колючих акул, долгохвлстов, лунников, гемпиловых и др., к этой мезобентопелагической группировке относ€тс€ и макропланктонные виды, близкородственные видам, характерным дл€ океанской мезопелагиали. ¬ их число вход€т, напомню, представители таких семейств, как топориковые (в том числе мавролик), фотихтовые, меланостомиевые, миктофовые, меламфаевые, меланоновые и др. ѕредполагалось, что основы ареалов этих макропланктонных видов наход€тс€ в системе горизонтальных и вертикальных вихрей, существующих над материковым склоном, но пр€мых доказательств этому нет. ќтсутствие у большинства таких рыб значительного выноса за пределы биотопа (вод, покрывающих склоны и подводные подн€ти€) на любых стади€х жизненного цикла свидетельствует о существовании специального механизма удержани€ их в присклоновой зоне, сущность которого остаетс€ пока непон€тной.
ќбщее число рыб, обитающих в мезобеитали и мезобентопелагиали, намного больше, чем в других океанических ихтиоцеиах. ѕо довольно приблизительному (но отнюдь не завышенному) подсчету (см. табл. 4), в этой зоне обитают около 2600 видов (почти 1500 без экотонпых шелъфо-склоновых рыб). ќбособленность этой фауны сильнее всего выражена, на видовом уровне, хот€ число родов, обнаруженных только в водах верхней части материкового склона и подводных подн€тий, также достаточно велико. ¬ то же врем€ к числу специфических мезобентальных таксонов более высокого уровн€ можно отнести только относительно немногие семейства, в то врем€ как остальные группы представлены либо па шельфе, либо в нижней части склона, а часто и в обоих биотопах.
¬ сообществах нижней части склона (батибенталь) и над ней (батибентопелагиаль) представлены сравнительно немногие систематические группы. »з хр€щевых рыб здесь живут только колючие акулы из родов Centropho rus, Centroscymnus, Somniosus, кошачьи акулы Apristurus, ромбовые скаты Bathyraja и Breviraja, а также носатые химеры-харриотты, по ни один вид не €вл€етс€ эндемичным дл€ биотопа. —реди костистых рыб наиболее характерны виды родов Aldrovandia, Synaphobranchus, Alepocephalus, Bathypterois, Bathysaurus, Antimora, Coryphaenoides, Cataetyx, Paraliparis, Lycenchelys и др. ѕо числу видов и особей в тропиках €вно доминируют три группы Ч гладкоголовы, долгохвосты и ошибневые, а в умеренных и холодных водах две Ч морские слизни и бельдюговые. —уммарное число видов достигает 450, но больша€ их часть (273 вида) оказываетс€ общей с нижней мезобенталью (граница двух сообществ батиали представл€ет собой экотонную зону, расположенную на глубине 1300Ч1800 м), а многие другие (91 вид) Ч с абиссобенталью, так что на долю специфической фауны приходитс€ менее 20% состава. ¬ нижней части склона различи€ между континентальной и талассной батибентальпыми группировками еще слабее, чем в мезобентали. ¬ »ндийском океане, например, по данным ё. ». —азонова, около 75% видов гладкоголовов встречаетс€ как на склоне континентов, так и на подводных подн€ти€х.
јбиссобентальные и абиссобептопелагическ€е ихтиоцены включают около 140 видов, однако эндемичные дл€ ложа океана виды очень немногочисленны. „исло специфических родов 10Ч12, среди них Ericara из гладкоголо-вых, Bathymicrops пз зеленоглазковых, Echinomacrurus пз долгохвостовых, Acanthonus и Typhlonns из ошибневых, Pachycara и Thermarces из бельдтоговых. ќстальные виды принадлежат к родам, представленным и в батибептали (Synaphobranchus, Einoctes, Bathytyphlops, CoryI haenoides).
ѕри изучении сообществ термальных источников, обнаруженных недавно в “ихом и јтлантическом океанах па глубине более 2000 м, вы€снилось, что в состав этих биоценозов вход€т и рыбы. ак показали подводные наблюдени€, фотосъемки и ловы, вблизи геотермальных изли€ний √алапагосского рифта встречено более 20 видов рыб (в основном макрурид и офидиид), но почти все они держатс€ в некотором удалении от мест вытекани€ теплой, насыщенной сульфидами и лишенной кислорода воды. ≈динственное исключение составл€ли особи одного вида, предварительно отнесенные к роду Diplacanthopoma (пока не удалось поймать ни одного экземпл€ра), которые по-одиночке, парами и тройками посто€нно по€вл€лись из кратера источника и снова уходили внутрь. ѕредполагаетс€, что эта рыба питаетс€ непосредственно бактери€ми, окисл€ющими сероводород. ¬ термальной воде живет также зоарцида Thermarces cerberus, €вл€юща€с€, по всей веро€тности, посто€нным членом этого своеобразного сообщества; другой представитель того же рода обнаружен только в периферической зоне рифтового Ђоазисаї. —лабовыраженный (только на родовом и видовом уровне) эндемизм гидротермальных рыб резко контрастирует с большой обособленностью бентосных беспозвоночных (подкласс погонофор, надсемейство моллюсков и т. д.).
¬ глубоководных желобах (хадобенталь) до сих пор обнаружено всего 7 видов рыб, ведущих дойный образ жизни (рис. 42). ¬ их числе три вида ошибневых Ч Leucicorus atlanticus (наибольша€ глубина вылова 6800 м в желобе ѕуэрто-–ико), Holcomycteronus profundissimus (7160 м в желобе —унда у южного побережь€ острова ява) и Abyssobrotula galatheae (8370 м в желобе ѕуэрто-–ико) и четыре вида морских слизней Ч Notoliparis macquariensis из желоба ћаккуори (глубина 5410 м), N. kurchatovi пз ёжно-—андвичева желоба (5470 м) (в табл. 4 эти два вида условно помещены в графу ј’), N. kermadecensis из желоба ермадек (6770 м) и Pseudoliparis amblysto-mopsis и неописанный вид этого же рода из урило- ам-чатского и японского желобов (максимальна€ глубина лова почти 7580 м). ¬се хадобентальные офидииды встречаютс€ и в абиссобентали (рекордист по глубине Ч абиссобротула Ч добывалась в восточной ѕацифике всего лишь на глубине 2330 м), тогда как оба рода липарид (они, кстати сказать, описаны ј. ѕ. јндри€шевым) эндемичны дл€ глубин более 5500Ч6000 м. Ќасколько сейчас известно, основную пищу всех этих видов составл€ют донные ракообразные (изоподы и амфиподы), но у кермадекских нотолипарисов, а их было поймано сразу 5 в одном трале, в желудках оказались также рыбь€ чешу€ и кости. ’ищничество представл€етс€, однако, очень маловеро€тным дл€ этих некрупных и к тому же в€лых и пассивных, суд€ по всем данным, существ. »х поймали скорее всего там, где они собрались дл€ поедани€ падали, опустившейс€ из верхних слоев океана. ¬ €ичниках двух самок этого вида было обнаружено 737 и 849 икринок, из которых самые крупные имели 7,5Ч9 мм в диаметре. ». Ќильсен предполагал, что этот вид, подобно батиабиссобентальному карепрокту Careproctus ovigerum, может вынашивать развивающуюс€ икру в ротовой полости. ћожно думать, что в хадобентали существуют в другие виды рыб, но их число вр€д ли может быть значительным.
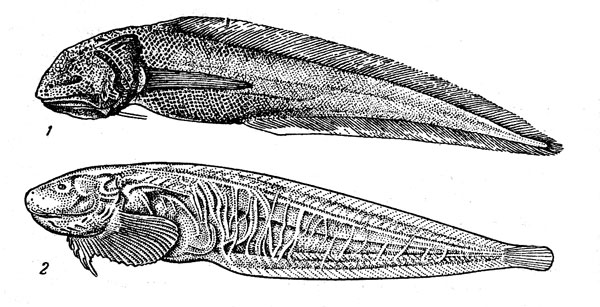
–ис. 42. –ыбы наибольших глубин абиссобротула (1) и псевдолипарис (2).
¬се ловы донных рыб в глубоководных желобах выполнены с помощью очень примитивною и, казалось бы, анахроничного оруди€ лова, которое и в наш век технического прогресса остаетс€ таким же, как в прошлом столетии. Ёто Ч трал —игсби, он же трал јгассица, названный так по фамили€м капитана американского корабл€ Ђјльбатросї и выдающегос€ зоолога, неоднократного руководител€ экспедиций на этом же судне. “рал этот предназначен дл€ сбора бентоса.
ѕредставьте себе две крутых дуги из железных пластин, скрепленных трубами и тросами, к которым прикреплен короткий мешок из грубой дели (внутри него помещаетс€ мелко€чейна€ вставка). Ёта снасть опускаетс€ па дно на длинном тросе и, будучи совершенно симметричной (у нее нет ни верха, ни низа), всегда занимает на грунте рабочее положение. —амо траление носит пассивный характер: судно, дрейфующее по ветру, медленно волочит трал, собирающий со дна все, что Ђплохо лежитї (точнее говор€, то, что лежит хорошо и не способно отплыть или отползти).
аждое траление в ультраабиссали - большое событие в экспедиционной жизни. ѕосадка трала —игсби па дно узкого желоба без применени€ специальной акустической аппаратуры (а она еще не нашла широкого применени€ в нашей практике) очень трудна. ќна требует точного расчета дрейфа судна, а также оценки необходимой длины вытравливаемого троса (эта оценка, производима€ Ђна глазїї с учетом двух переменных - глубины по эхолоту и угла наклона троса, уход€щего в воду,Ч самое слабое звено методики работ). Ѕывает, что трал не достигает дна или. наоборот, лишний трос завиваетс€ в кольца, Ђколышетс€ї и рветс€ при
выборке, так что долгие часы работы пропадают впустую. ћне приходилось присутствовать при нескольких дес€тках удачных тралений в самых разных желобах (в том числе и в самом глубоком из них - ћарианском), по только дважды - в японском и ёжно-—андвичевом желобах они приносили рыб. ясно, что на хадальных глубинах их совсем не так уж много.
Ёкологи€ донно-придонных рыб открытого океана исследована, пожалуй, еще хуже, чем экологи€ рыб глубоководной пелагиали. ƒанных о их размножении, росте, питании и других биологических особенност€х пока немного, и обща€ картина, складывающа€с€ из разрозненных фрагментов, не может считатьс€ вполне адекватной: многие вопросы еще ждут ответа. “ем пе менее уже сейчас вы€влены некоторые общие закономерности, характерные дл€ рыб бентопелагического комплекса.
–азмножение. ’р€щевые рыбы, обитающие в придонных сло€х, демонстрируют все три возможных в этой группе типа репродукции Ч €йцерождение, €йцеживорождение и живорождение. числу €йцеживород€щих принадлежит п латимери€, но из костистых рыб этим путем размножаютс€ только некоторые эпимезобентальные морские окуни (Sebastes) представители семейств Bythitidae и Aphyonidae. ѕреобладающее большинство видов имеет пелагические икринки, число которых сильно варьирует: от нескольких тыс€ч (у аргентин и батитифлопсов) до сотен тыс€ч (у моровых, долгохвостов, крупных камбаловых). ” рыб, откладывающих донную икру (икру с отрицательной плавучестью), плодовитость, как правило, заметно меньше и измер€етс€ дес€тками Ч сотн€ми икринок (минимально Ч 20 икринок у Rinoctes nasutns), по у некоторых видов гладкоголовов (крупные представители рода AlepocepLalus) может достигать 10 тыс.
—уд€ по встречаемости личинок и мальков в уловах, донно-придонных рыб можно разделить на две группы. ” одних постэмбриональное развитие проходит в толще воды, у других Ч в придонных сло€х. первой группе, дл€ которой характерны значительные морфологические преобразовани€ в конце личиночно-малькового периода (метаморфоз), должны быть отнесены все угри и родственные им галозавриды и нотакантиды, также имеющие лептоцефальную стадию, некоторые хлорофтальмиды (Bathytyphlops с мальком макристиеллой) и сиподонтиды (Balhysaurus с мальком макристиумом), те офидииды, которые обладают специфическими личиночными стади€ми типа Ђэксериллиумї или Ђвексиллиферї, а также, по-видимому, мориды и долгохвосты. ¬торую группу образуют алепоцефалиды, хлорофтальмиды из родов Ipnops и Bathymicrops, некоторые офидииды, все липариды и зоарциды и др.
Ёта проста€ классификаци€ не может быть распространена, однако, на всех рыб бентопелагического комплекса, поскольку у многих из них ранние стадии развити€ все еще остаютс€ неизвестными. ќна не €вл€етс€ бесспорной даже в отношении некоторых из перечисленных выше рыб. ѕредставлени€м о наличии пелагической стадии у всех видов семейств Moridae и Macrouridae противоречит, например, существование в этих группах локального эндемизма: в обоих семействах сейчас известно немало видов, которые ограничены в своем распространении преп€тстви€ми, непреодолимыми дл€ них, казалось бы, только во взрослом состо€нии Ч дл€ батибентальных рыб это мелководные пороги, отдел€ющие глубинные бассейны, дл€ мезобентальных Ч океанические глубины, окружающие подводные возвышенности. Ћичиночно-мальковые стадии всех массовых доино-придопных рыб, особенно насел€ющих батиаль, вообще очень редки в толще воды и почти никогда не встречаютс€ вдали от материковых склонов и, как уже говорилось, горных подн€тий в открытых водах.
¬ св€зи с этим высказывались предположени€ о том, что эмбриональное и раннее постэмбриональное развитие таких рыб может проходить в придонных горизонтах.
–азмеры и рост. ¬ среднем донно-придонные глубоководные виды рыб крупнее пелагических глубоководных, но мельче тех, которые живут в верхней толще воды (см. рис. 27). —реди них нет таких гигантов, как эпипелагические акулы, марлины и меч-рыбы, но некоторые из батиальных видов достигают все же значительной длины. числу наиболее крупных относ€тс€ пол€рна€ акула (длина 6,4 м при массе около 1 т), шестижаберна€ акула Ќехаnchus griseus (4,8 м), акула-домовой (3,3 м), ложнокунь€ акула (3 м), звездчатошипа€ акула Echinorhinus brucus (2 м), ромбовые скаты рода Bathyraja (2 м), химеры 1(1,5 м), а также латимери€ (длина 2 м, масса почти 100 кг). »з костистых рыб наибольших размеров достигают белокорый палтус (длина до 4,7 м и масса более 300 кг), клыкач (2,1 м), некоторые долгохвосты, ателеоиовые и гемпилиды (по 2 м), слитиожаберные угри (1,8 м), зубатки (1,4 м), иекоторые офпдииды (до 1,3 м), а самыми маленькими €вл€ютс€ бентопелагические стерпоптихиды и миктофиды из родов Maurolicus, Polyipmis, Diaphus, Laitipanyciodos, а также бептические липариды и перцихтиды из родов Paraliparis, Osopsaron, Pteropsaron, длина которых не превышает 5Ч6 см. ¬ абиссали размерный спектр значительно уже: от 19Ч20 см у некоторых гладкоголовов (Rmoctes, Mirognathus) до 85 см (при массе 7 кг) у батизавра Bathysaurus mollis, который, как показано пр€мыми наблюдени€ми с подводного аппарата, способен к резким рывкам на рассто€ние 10Ч15 м и, без сомнени€, должен быть отнесен к нектону (в толще воды нектонные рыбы отсутствуют в батипелагиали и ниже).
√лубоководные донные и придонные рыбы, как правило, отличаютс€ медленным ростом и большой продолжительностью жизни. »сключение составл€ют, пожалуй, только мелкие беитопелагические виды (мавролик, свет€щиес€ анчоусы) из семейств, характерных дл€ открытой пелагиали, которые совершают суточные вертикальные миграции из придонных горизонтов в эпипелагиаль. ¬се они, подобно их чисто пелагическим родичам, созревают в возрасте, близком к годовалому, и живут ненамного больше. ” таких рыб, как колючие акулы, аргентины, гладкоголовы, моровые, крупные макруриды (род Coryphaenoides), большеголовы, скорпеновые, камбалы, продолжительность жизни варьирует, насколько сейчас известно, от 10 до 30 и даже 40 лет, причем годовые приросты длины редко превышают 3Ч5 см. ѕоловозрелыми опи станов€тс€ в возрасте 5Ч10 лет, а иногда € позже. Ёти рыбы с полным правом могут быть отнесены к категории длинноцикловых.
ѕитание. ѕредставл€етс€ естественным, что рыбы, живущие на дне или у дна, должны питатьс€ в основном пищей, наход€щейс€ на грунте, т. е. бентосными организмами. Ёто вполне логичное заключение, которое, как правило, оправдываетс€ в приложении к донпо-придонным рыбам шельфовой зоны, оказываетс€, однако, совсем не бесспорным, когда речь идет о глубоководных видах Ч почти все массовые формы потребл€ют в пищу пелагических животных (мезо- и макропланктон, микронектон). “ака€ Ђдиетаї характерна не только дл€ рыб, поднимающихс€ в водную толщу, но и дл€ многих посто€нных обитателей дна.
«начительна€ часть рыб бентопелагического комплекса относитс€ к числу хищников, поедающих крупную добычу Ч других рыб, головоногих, ракообразных. Ќаиболее глубоководными из донных хищников €вл€ютс€ батизавры, в пище которых зарегистрированы слитножабериые угри, макруриды, лиценхелы, креветки, кальмары. »звестен и способ их охоты: рыбы поджидают добычу, полусто€ или лежа на дне в слегка наклонном положении (передн€€ часть тела приподн€та), и лов€т ее неожиданным стремительным броском, сопровождаемым хватательным движением челюстей. Ќекоторые хищники мезобентопелагиалн способны к совместному добыванию добычи. “ак поступают, к примеру, некоторые колючие акулы, а также рексии. Ёти рыбы режут крупную рыбу на куски своими мощными зубами, а затем заглатывают по част€м, и части одной жертвы иногда обнаруживаютс€ в желудках разных хищников, пойманных одним тралом.
онечно, на тех участках дна, где имеетс€ достаточное количество кормового бентоса, он также идет в пищу рыбам. ћногие из преимущественно планкто€дных видов, в частности гладкоголовы, долгохвосты, мориды, нередко разнообраз€т свое питание за счет донных моллюсков, полихет, ракообразных. ак показал –. ’эдрич, отдельные группы видов (подроды) рода Coryphaenoides, различающиес€ размерами и строением зубного аппарата, различаютс€ и по характеру пищи. ћелкие виды со слаборазвитым зубным вооружением (например, —. carapinus) поедают главным образом бентос, в первую очередь офиур, тогда как более крупные виды с сильными зубами Ч пелагических ракообразных, рыб, головоногих. ѕреимущественно бенто€дными €вл€ютс€ многие офидииды, угри, камбалы, чисто бенто€дными Ч галозавры, спиношипы, линариды, встречающиес€ в св€зи с этим только на м€гких грунтах.
»нтересно сопоставить трофическую структуру глубоководного донного ихтиоцена и таксоцена беспозвоночных того же сообщества. Ѕольша€ часть глубоководного макробентоса, по ћ. Ќ. —околовой, входит в состав пищевой группировки детритоедов, питающихс€ органическим веществом донных осадков. “акже довольно многочисленны среди донных беспозвоночных сестоноеды, пищу которых составл€ют взвешенные в воде органические частицы, включа€ живой планктон. ѕлото€дные беспозвоночные, потребл€ющие живую добычу и трупы, представл€ют меньшинство и в сессильвом (сид€чем) и в вагильном бентосе, но решительно преобладают в нектобентосе, (это в основном придонные ракообразные). —реди рыб чистых детритоедов, по-видимому, нет, хот€ остатки грунта, заглоченные, можно думать, вместе с другой пищей, не раз обнаруживались в желудках бентофагов (например, у спиношипов и галозавров). числу сестонофагов-хватателей (почти что сессильных) можно отнести батиптеров, возвышающихс€ над дном на треножнике, составленном лучами хвостового и брюшных плавников, а возможно, и других глубоководных хлорофтальмид (кроме ипнопсов). ѕреобладают в ихтиоцене, конечно, плото€дные, среди которых следует выдел€ть потребителей донных беспозвоночных, все€дных хищнп-ков-падалыциков и насто€щих хищников, охот€щихс€ только на крупную живую добычу.
¬ершинные участки подводных подн€тий, как правило, имеют скальное дно, полностью лишенное рыхлых осадков, что само по себе определ€ет доминирование рыб-планктофагов в сообществах таких биотопов. ƒо самого недавнего времени бытовали два взгл€да на источник пищевых ресурсов, определ€ющих существование и продукцию Ђгорныхї попул€ций рыб. —огласно одному из них, продуктивность подн€тий определ€етс€ локальным орографическим эффектом Ч подъемом глубинных вод, обусловленным завихрением набегающего на гору потока посто€нного течени€ и привод€щим в действие механизм биологической продуктивности в его полном цикле: от повышени€ концентрации биогенных солей в эвфотическом слое к увеличению первичной продукции водорослей и за ее счет к увеличению биомассы зоопланктона. ƒл€ более точного (Ђболее наукообразногої) объ€снени€ этого €влени€ была выдвинута гипотеза о формировании над вершинами квазистационарных вихрей, которые п обеспечивают посто€нный подъем вод, будучи настолько изолированными от окружающих вод, что внутри них образуютс€ самосто€тельные экологические системы. ƒействительно, подводный рельеф, усложн€€ в известной степени структуру набегающего потока, может способствовать образованию локальных вихрей, привод€щих в конечном счете к обогащению поверхностных слоев биогенными элементами. ќднако пространственно-временна€ устойчивость таких впхрей представл€етс€ довольно слабой, а их продолжительна€ изол€ци€ Ч в высшей степени проблематичной. ¬едь врем€, необходимое дл€ развити€ фитопланктона, исчисл€етс€ несколькими сутками, а кормова€ база рыб Ч мезопланктон и макропланктон Ч развиваетс€ намного дольше, соответственно около мес€ца и полугода.. —толь длительное существование над подводной возвышенностью неподвижных и существенно замкнутых вихревых образований почти неверо€тно: если вли€ние локального обогащени€ поверхностных вод биогенами и может отразитьс€ на продуктивности зоопланктона и тем более па рыбопродуктивности, то произойдет это только в значительном удалении от места подъема вод. “аким образом, эффект увеличени€ биомассы кормовых организмов за счет локального Ђапвеллингаї может быть использован не бептопелагическими рыбами той подводной возвышенности, котора€ послужила причиной возникновени€ такого Ђшлейфовогої п€тна повышенной биомассы планктона, а только истинно пелагическими хищниками (например, тунцами и кальмарами). ѕоэтому гораздо более правомочным представл€етс€ другой подход, св€зывающий кормность подн€тий с горизонтальным приносом (адвекцией) пищевых объектов набегающим потоком. ѕр€мые исследовани€ питани€ рыб, живущих над подводными возвышенност€ми, показывают, что их откорм происходит в основном за счет макропланктона, вход€щего в состав мигрирующих звукорассеивающих слоев (миктофиды, эвфаузииды и т. п.) и опускающегос€ днем на самую вершину горы (если, конечно, она лежит выше нормальной глубины дневного опускани€ этих слоев). –аспространение. √еографическое распространение океанических рыб, вход€щих в состав бентопелагического комплекса, еще не изучено в детал€х, но представл€етс€, что размеры их ареалов сильно варьируют. Ќекоторые из них весьма обширны Ч такие виды, как, например, эпимезобентальные антигони€ Antigonia capros и рыба-бекас Macrorhamphosus scolopax, мезобентальпые м€гкоголовый долгохвост Malacocephalus laevis и низкотелый берикс ¬егух splendens, мезобатибеитальные альдрованди€ Aldrovandia phalacra и антимора Antimora roslrata, батибентальные гладкоголов нарцет Narcetes sloimias и ксиелациба Xyelacyba myersi, батиабиссобентальные гладкоголов беллоци€ Bellocia koefoedi и батизавр Bathysaurus mollis, абиссобентальные слепой тифлонус Typhlonus nasus и эхиномакрурус Echinomacrurus mollis, обнаружены на соответствующих глубинах в тропических водах всех трех океанов. ¬ то же врем€ во всех вертикальных зонах есть и виды, которые известны из очень ограниченных участков и, по-видимому, на самом деле имеют очень узкое распространение (хот€ представлени€ об эндемизме могут основыватьс€ на недостаточных данных). райний случай почти Ђточечногої ареала демонстрирует распространение латимерии, обитающей только на склонах двух маленьках островов оморского архипелага. Ѕольшинство других узколокальных эндемиков также обитает на вершинах подводных подн€тий.
»звестно, что дробность биогеографического делени€ бентали, достигающа€ наибольшей выраженности на шельфе и в верхней части склона (т. е. на глубинах залегани€ поверхностных и подповерхностных вод), уменьшаетс€ в нижней батиали и сводитс€ к минимуму на ложе океана, где можно обнаружить только самые главные границы (те, которые раздел€ют регионы высшего ранга Ч области). ћногими работами показано, что главные зоогеографические границы в мезобентали как бы продолжают вглубь границы, проход€щие на шельфе. ƒействительно, анализ географического распространени€ рыб, насел€ющих склоны континентов и подводные подн€ти€, дает возможность вы€вить те же крупномасштабные регионы, что и в сублиторальной зоне. “ак, √. ј. √о-ловань, рассматрива€ ареалы мезобентальных рыб, обитающих у западных побережий јфрики и Eвропы, обнаружил в составе ихтиофауны 12 географических элементов и выделил четыре района Ч бореальный, северный субтропический, тропический и южный субтропический Ч в районировании шельфа.
Ќемногочисленность поимок многих видов батибентальных рыб сильно затрудн€ет анализ их географического распространени€. “ем не менее представлени€ о преобладании в этой зоне узколокальных ареалов, существовавшие до недавнего времени, не подкрепл€ютс€, как правило, новыми данными. ѕочти дл€ всех видов, известных по многим нахождени€м, характерно широкое распространение. ƒостаточно четкое разделение существует, однако, между фаунами тропических, умеренных и холодных вод. ¬ тропической зоне выдел€ютс€ экваториальный и широкотропический типы ареалов с циркумглобальным, индовестпацифическим, атлантическим и восточно-тихоокеанским подтипами. ”меренноводные ареалы подраздел€ютс€ на потальные и бореальные, а холодноводные Ч на антарктические и арктические.
„то касаетс€ абиссобентальных и хадальных рыб, все они известны по немногим, часто единичным находкам, что сильно затрудн€ет рассмотрение их зоогеографии. Ќужно отметить, впрочем, что на ложе океана некоторые виды уже сейчас найдены в далеко разобщенных районах (в двух или трех океанах встречены Bathymicrops regis, Ipnops meadi, Holcomycteronus profundissimus, Abyssobrotula galatheae и др.), и это позвол€ет предполагать их широкое распространение.
|
ѕќ»— :
|
ѕри использовании материалов сайта активна€ ссылка об€зательна:
http://udimribu.ru/ '–ыбалка'